Block Diagram
In an old-fashioned, block-diagram view of the brain, the blocks around the outside might have labels like “visual cortex,” “auditory cortex,” “motor cortex,” and so on. Implicitly, these are presumed to work like the peripherals of a computer, carrying out specialized processing to deal with their particular modality. (Robots, too, are usually built this way.)
Neural signals would presumably flow in the obvious direction: inward for sensory modalities, outward for motor. Convolutional neural nets, whose design was directly inspired by visual cortex, seem in keeping with this view. They are fully feedforward, meaning their connectivity flows one way, from a “retinal” input layer to semantically meaningful output in an embedding layer.
Those semantic embeddings, in turn, seem like the right kind of input to … what? Something needs to go in the middle, taking in the processed sensory input, making decisions, and generating high-level motor commands; we might as well call this the Central Processing Unit, though the usual innocuous label is something like “association cortex.” It’s hard to look at that block on the diagram and not think “homunculus.” For surely, that’s where “you” live. A perceptron, which merely turns pixels into embeddings, does not seem like a promising spot in the diagram for locating anything akin to a “self.”
However, as the motor-first perspective laid out in chapter 4 implies, almost everything about this block diagram and its information-processing paradigm may be wrong. For one, I’ve argued that there is no homunculus. If there were, unplugging it would surely turn the lights out. Instead, when a good portion of the non-sensory-specific “association cortex” in the frontal lobes is destroyed or disconnected from the rest of the brain—as happened to many victims of the mid-twentieth-century lobotomy craze—the effects can be disconcertingly subtle, just as with split-brain patients.
Emphasis is warranted on can be. The details of the procedure, and the nature and extent of the ensuing damage to the brain, varied considerably. Some victims of this brutal medical fad were killed outright by massive cerebral hemorrhage, or rendered profoundly disabled. After her lobotomy in 1941, for instance, Rosemary Kennedy, President John F. Kennedy’s sister, was left unable to walk, speak intelligibly, or use the bathroom; she required constant care throughout the rest of her life.
Nonetheless, in 1942, star neurosurgeons Walter Freeman and James Watts claimed that, of the two hundred lobotomies they had performed to that point, sixty-three percent were “improved,” twenty-three percent were “unchanged,” and only fourteen percent were “worse.” 1 Rosemary Kennedy was one of those two hundred, and was presumably counted among the “worse.” While we must meet with skepticism any assessment of what constituted an “improvement,” figures like these do make it clear that most lobotomized patients didn’t end up like her. What, then, wondered Freeman and Watts, were the frontal lobes even for?
Scene from a 1944 film extolling the virtues of prefrontal lobotomy for the treatment of schizophrenia
“Neurologic manifestations are surprisingly slight following prefrontal lobotomy, and the old concepts of the frontal lobes as concerned with the higher integration of movements have had to be revised. […] The psychiatrist must think out these problems and determine for himself what contribution the frontal lobes make to normal social existence […].” 2
Another puzzle: as described in chapter 5, cortex is highly modular. In particular, while subtle differences exist between sensory and motor cortex, their structure and wiring are surprisingly similar. Both are dominated by recurrent connections, or feedback—shades of Wiener’s cybernetics.
That seems especially odd for sensory areas. Perceptrons are entirely feed-forward, and they do seem to capture something of what is going on in visual cortex. The “receptive fields” of units early in a convolutional net (that is, the visual patterns to which these model neurons are most sensitive) look uncannily like the receptive fields of real neurons early in the visual system. The sparse, semantically meaningful embeddings in the final layers of perceptrons also resemble the activation patterns recorded from higher-level visual neurons. 3
Observations like these have led to debates between neuroscientists and AI researchers. Some (especially on the AI side) emphasize the functional similarities between brains and deep learning. Neuroscientists, at times irritated by the looseness of these parallels, point out the inconsistencies, including:
- Real neurons communicate with spiking action potentials, not the continuous values of artificial neural nets.
- Neurons and synapses are vastly more complex than their cartoonish deep-learning versions.
- The backpropagation algorithm used to train perceptrons isn’t biologically plausible. 4
- Deep learning, true to its name, often relies on dozens or even hundreds of layers, while the brain only has a few layers. Brains would be useless if visual stimuli needed to pass through so many cortical layers, because neurons and synapses are so slow relative to computers; in real life, there’s often only time for sensory input to propagate through a few synapses before it must become a motor output, driving a behavioral response. And, of course,
- Visual cortex, like all cortex, is dominated by feedback connections, while perceptrons are entirely feed-forward.
The arguments have gone back and forth. Some of the objections raised by neuroscientists are easy to counter. For instance, the universal function-approximation theorems mentioned in chapter 3 prove that, in principle, networks of complex spiking neurons can be approximated by (larger) networks of highly simplified model neurons. Researchers have also come up with versions of backpropagation and related learning rules that are more biologically plausible, and, while we don’t yet fully understand the neuroscience, there is evidence of such learning rules in the brain. 5
In 2018, machine learning researchers found that convolutional neural nets could reliably be fooled into making bizarre misclassifications using “adversarial stimuli,” which many at the time considered conclusive evidence that deep learning wasn’t anything like biological vision. 6 But countervailing findings in 2022 showed that the primate visual system could be fooled in just the same way, and to the same degree! 7 So, the debate continues.
Clearly, the shallowness of visual cortex and its highly recurrent connectivity can’t be finessed—in these respects, the brain is self-evidently not like a convolutional neural net (CNN). Structurally, visual cortex—in fact, all cortex—looks more like a “recurrent neural network” (RNN).
Recurrence
As the name implies, recurrent neural networks have both forward and backward connections. More formally, this means that neurons take inputs not only from other neurons in space (i.e., a previous layer) but also in time. That is, the output of a neuron at time step t becomes an input for neurons at time step t+1; weights associated with these inputs define the recurrent part of the network’s connectivity.
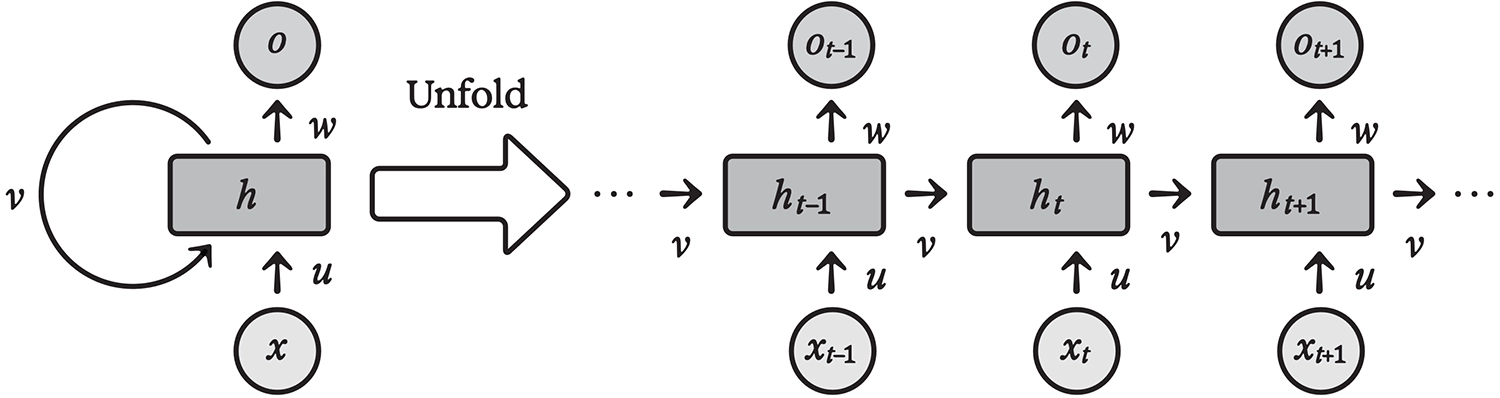
Architecture of a recurrent neural network, shown to the right “unfolded” over time steps t–1, t, t+1, ...
Keep in mind that feedforward artificial neural networks are, by definition, both timeless and memoryless. When they evaluate an input, they do so independently of any other input; no time variable is involved. By introducing time, RNNs extend neural computation into the dynamical realm. That’s more realistic, as any physical system (including the brain) unfolds over time. The input weight associating each neuron with its own activation in the previous time step implements a memory mechanism in its simplest form: a degree of persistence in neural activity.
Recurrent networks are generalizations of feedforward networks, in the sense that any feedforward net, like a CNN, can be implemented as a recurrent net. To see how, imagine replacing the CNN’s “spatial” feedforward connections with equivalent feedback connections. A CNN with one hundred layers would turn into an equivalent RNN that runs in one hundred time steps.
Seen this way, the high proportion of recurrent connections in visual cortex might seem less surprising; those recurrent circuits could be implementing something like deep learning, but doing so in time rather than in space. In fact, since every kind of sequential neural processing must happen over time, the simple remapping envisioned here amounts to little more than notational sleight of hand.
But what about reaction time? It would hardly be a good idea for you, as, say, a highly visual prey animal, to experience the world a hundred time steps in the past. 8 At t=0, a tiger might just be coming into view through the bushes. At t=8, it might have caught sight of you. At t=16, it might be tensing to pounce. By t=80, you might already be getting eaten, so there will be no t=100!
No problem. Even when deep convolutional nets are optimized purely for accuracy, researchers have found it beneficial to add “skip connections” to the standard sequential information flow, allowing some activations to bypass layers. 9 Moreover, while making a feedforward architecture deep is a way of improving task performance, “early exits” can be used if the extra accuracy is unneeded—or if reaction time matters.
After just a few CNN layers, or RNN time steps, the network can make a reasonable guess as to whether a cat is in the picture. Since the network’s accuracy isn’t great at this point, it can be tuned to prefer false positives over false negatives and immediately escalate the timely alert. That is indeed how we operate for highly salient stimuli; hence, the double take. (What’s that, a cat?)
A few more layers, or time steps, can accurately establish whether the cat in question is of the small Felis domesticus kind or the potentially man-eating Panthera tigris. (Whew! It’s just a house cat.) It may take dozens of further layers, or time steps, to pinpoint the breed, guess the color of the eyes, establish its mood and likely state of mind; but in the recurrent setting, this is where we would say “upon closer examination….” Recurrent nets, then, can implement both rapid responses and the many-layered precision of CNNs, operating dynamically rather than as static functions.
This dynamic aspect is the heart of the matter. The visual world is not a succession of uncorrelated still images, and the visual cortex is not an abstract, timeless mathematical function. In a continuously changing and temporally correlated world, where one’s actions (including perceptual actions, like saccades) determine what one sees, highly recurrent architectures are to be expected and classification is better thought of as continual prediction.
Evaluating a CNN-like function from scratch for every visual frame would be madness; for one, it would be computationally wasteful, since, most of the time, each frame is nearly the same as the previous one. Worse, independent frame-by-frame processing would be incapable of reconstructing the visual world as it actually is. Building and maintaining the “controlled hallucination” described in chapter 4 requires integrating information over many frames. And that means memory. It is, almost by definition, a job for a recurrent network with persistent state.
Demonstrations of Portia gaze tracking from the lab of Dr. Elizabeth M. Jakob, UMass Amherst
Imagine how the visual system of a Portia spider might work. Owing to the very narrow field of view of the spider’s high-resolution front-facing eyes, a frame-by-frame CNN would be nearly useless; for Portia, understanding a visual scene requires moving the eyes around dynamically and reconstructing a model of the world over time—not unlike the way a blind person can “see” someone’s face by touch, using their fingertips, though in Portia’s case, it would be more like a single fingertip. Large predators like birds, frogs, and mantises are the bane of Portia spiders, seemingly because they are too big to recognize in time! 10
Humans can see more at once than Portia, but not nearly as much as we believe. If we were to experience our visual input feed in a rawer, less “hallucinated” form, it would look something like the shaky, grainy found footage of a horror film like The Blair Witch Project, with a flashlight beam jumping around spasmodically to illuminate a tree branch here, a bit of a face over there, the corner of a woodshed, a dark something on the ground. That horror trope will give you perceptual claustrophobia, a sense that no matter where you look, the stuff you really need to know about is happening offscreen—to one side, or above, or behind your back. Like Portia being hunted by a giant mantis, we would wish we could zoom way out and see what the hell is going on!
From The Blair Witch Project, 1999
If you don’t normally feel that Blair Witch sensation of near-constant panic—and dear reader, I hope you don’t—it’s not because you can see so much more at a time. It’s because your controlled hallucination is good enough to make you feel that you do see everything happening “offscreen,” although you can’t. That is, you have confidence that your continually updated prediction is accurately modeling every behaviorally relevant feature of your environment, well beyond the narrow cone of that jittery foveal flashlight beam.
Efference Copy
Prediction underlies intelligence at every scale. Per chapter 2, single cells like bacteria must predict sequences of events, both internal and external, to survive. Emerging evidence suggests that it’s also what single neurons do; their synaptic learning rules appear to give rise to local sequence prediction. 11
Cortical circuitry, too, appears to implement predictive sequence modeling. 12 This may account for the highly recurrent architecture of cortex, whether in perceptual, motor, or so-called “association” areas: since sequences unfold over time, any sequence predictor must also operate in time, which requires feedback connections.
In visual (and other perceptual) areas, sequence prediction will also involve recognizing hierarchies of increasingly invariant features, like those of a feedforward CNN trained using the masked-autoencoder approach. This is because invariant features can be thought of as predictions. If you recognize that you’re looking at a banana, for instance, you can predict that it will remain a banana in the future, even if it rotates, you move closer or farther from it, or you look away and then look back at it.
Now, consider what happens when multiple brain areas are connected, entangled in mutual prediction. Actually, we already have. Several brain areas are involved in eye movement, 13 but let’s ignore the anatomical details and pretend a single “eye motor region” sends signals to the small, fast-acting muscles that aim the eyes.
First, consider the motor region in isolation. To carry out its own predictive processing, it will need to learn the basics of controlling eye movement: how to send signals to the muscles that result in predictable responses by the stretch receptors in those muscles. Three angular variables define each eye’s position, but six muscles control the movement, and many individual nerve fibers innervate each one. Our developmental programming ensures basic routing, such that relevant sensory and motor signals wire up to neurons in the right general areas. Still, learning oculomotor control is not trivial. When the sensorimotor loop isn’t perfectly tuned, the results may manifest as “lazy eye” or the oscillating pattern known as “nystagmus.” Wiener would recognize these as symptoms of excessive or insufficient negative feedback; the latter is akin to intention tremor.
Nystagmus
Meanwhile, the visual cortex will try to predict what imagery the retinas will see at any given time. In the service of that prediction, it will reconstruct an entire three-dimensional world. This seems so much loftier than what the eye motor region purportedly does that we might now be tempted to return to the homuncular fallacy, but with the shoe on the other foot: imagining that the visual cortex is the “boss” or “central processing unit” that orders the eye motor region around, telling it where to look next, treating it as a mere “peripheral.”
But the shoes are, so to speak, on both feet. The eye motor region can and does move the eyes around of its own accord, perhaps in response to a vestibular input, a sound, a touch, or something else. It could even do so in response to nothing. After all, we decide to do things all the time for reasons of our own, that is, not as immediate responses to an outside stimulus. Remember that, in a distributed picture of self, these decisions could be initiated by nearly any brain region. All of your brain regions are “you.”
Communication between the visual cortex and the eye motor region, then, does not merely consist of a one-way stream of gaze coordinates sent from one region to the other. It’s a two-way dialogue, communicating (in compressed form) the most useful aspects of the state of each region to the other.
By most useful, I mean most helpful in predicting the future for the receiving region. In general, rather than imagining region A sending “commands” to region B, imagine region B containing “sensory” neurons in region A, with the goal of finding the most salient information for making its own predictions. Visual cortex, then, will want to learn from the eye motor region about eye movements when they occur, because that will help predict what the visual cortex “expects” to see next. The eye motor region, in turn, will want to learn from visual cortex where the most interesting spots in the environment are, because knowing those will help predict where the eyes will next look. The regions are cooperating to better predict themselves, and in the process, better predict each other. Yes, it sounds more than a bit like theory of mind—but carried out between brain regions!
This picture makes a counterintuitive experimental prediction: that signals specifying movement will actually be sent from motor regions to other parts of the brain, rather than vice versa. And that is indeed the case: it’s called the “efference copy.”
In the mid-nineteenth century, pioneering German physicist and proto-neuroscientist Hermann von Helmholtz first suggested that something like the efference copy must exist. He noticed that if you cover one eye, then press gently on the other eyeball (but do this through the lid please, and gently—don’t hurt yourself and then blame me) you will see the world appear to move. This is interesting, because when you move your eyeball the usual way, the world seems to stay rock steady, although the images your eyes see are anything but steady. The visual system must therefore do something like self-motion cancellation, and this must make use of an extremely accurate real-time eye movement signal—the efference copy. Helmholtz reasoned that when you press your eyeball, there is no efference copy from the eye muscles corresponding to that displacement, so the visual system’s usual “motion cancellation” doesn’t compensate for it.
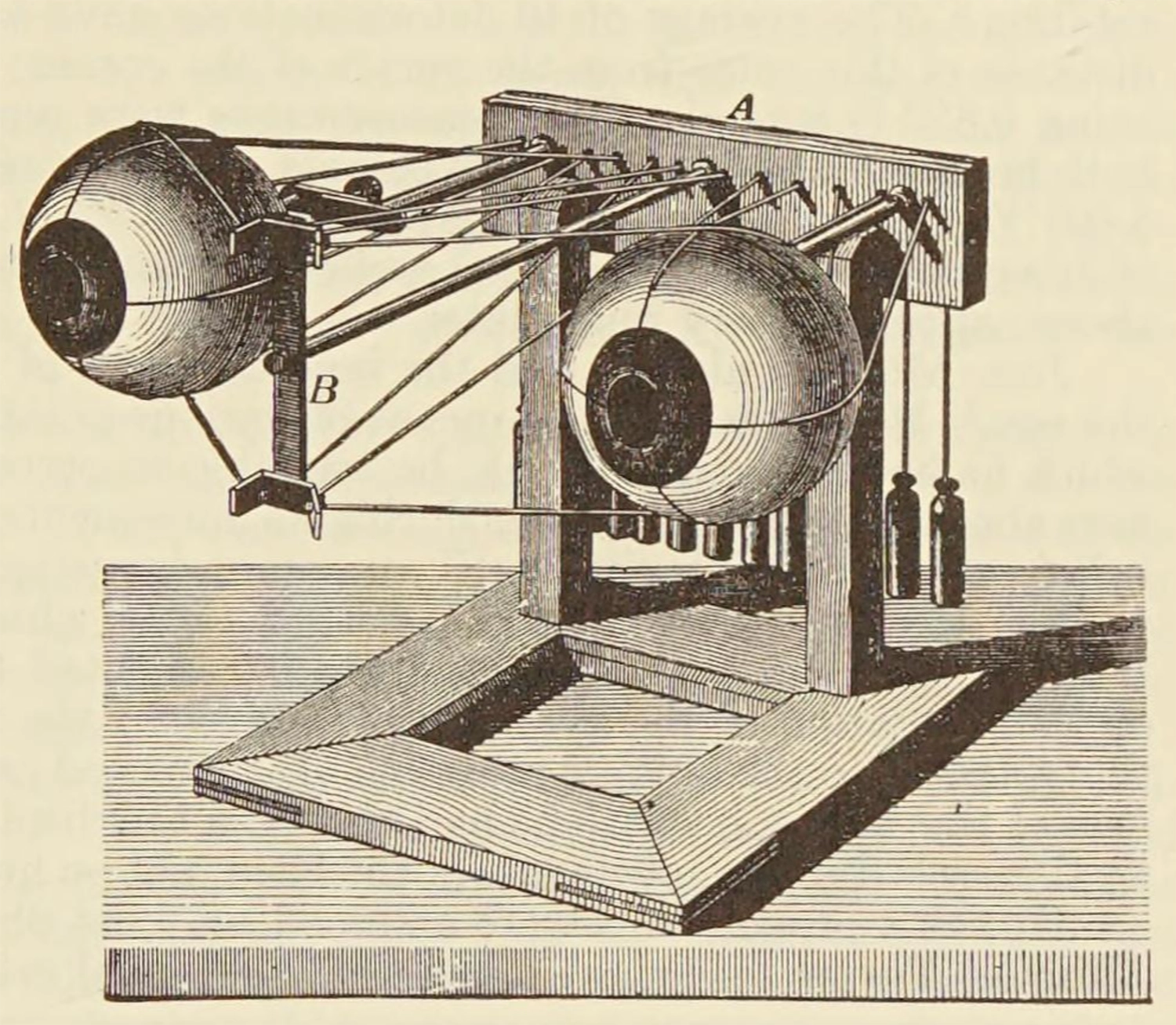
A mechanical model of ocular control, from Helmholtz’s Treatise on Physiological Optics
Helmholtz was right, but in 1900, Sir Charles Sherrington, an enormously influential neuroscientist who later won the Nobel Prize, cast doubt on the efference copy idea, causing it to languish for decades. 14 Having spent a significant part of his career exploring spinal cord reflexes, the sensorimotor pathways Sherrington had mapped out didn’t seem to involve anything like an efference copy, just stimulus, response, and feedback from stretch receptors in the muscles. 15 Later in the twentieth century, though, as neural recording technology improved, numerous studies found that efference copies are real and ubiquitous—not just for eye movements, but for all motor activity, and across species. 16
But why is it called a “copy”? Perhaps because the homuncular fallacy dies hard; we persist in imagining that motor regions are sent movement “commands” from on high, and then (somewhat bafflingly) send a carbon copy of these “commands” back to the sender—presumably, the “you” part. In reality, your motor regions are as much “you” as any other part of your brain, and while signals traveling to the motor regions surely carry information about future movement, thinking of them as “commands” and of signals going the other way as “copies” of those commands misses the point.
In the case of eye movement, the mistake may be easy to make because the eyes do generally look at whatever is most interesting, but that neither makes the motor region subservient to the visual cortex nor alters the fact that the motor region and the visual cortex are both doing the same thing: prediction.
Phenomenality
In case you’re not yet convinced that the brain serves the muscles at least as much as the other way round, consider the meaning of “behaviorally relevant,” the term I’ve used to describe what is in our umwelt, that is, what we perceive and what we care about. Behaviorally relevant means relevant to the muscles! If we can’t act on it, it doesn’t matter. And for creatures like us, just as for early bilaterians like Acoela, all action is muscular.
To be sure, a long evolutionary road leads from worms to humans … but it’s a continuous road. Our bodies are still, to no small degree, autonomous entities, surprisingly independent of our more recently evolved brains. Remember, if kept alive, our hearts will keep beating even if removed from our bodies, as Indiana Jones found out in the Temple of Doom. Like an inner worm, our gut continues to work under largely local control, with a decentralized “autonomic” nervous system serving the original functions of long-range muscular coordination.
Our blood vessels, too, are made of muscle, and regulate the flow of nutrients to every region of the body. To put it bluntly, they decide who gets to eat, and when. Because the brain is so voraciously energy-hungry, regulation there is especially fine-grained, with individual capillaries feeding less than a cubic millimeter of brain tissue dilating and contracting from second to second. This differential flow of oxygenated blood is the signal measured by functional magnetic resonance imaging (fMRI), one of our most important tools for mapping brain activity.
The brain, in turn, consists of multiple systems layered atop each other, repeating the same broad functional pattern: newer or “higher” levels provide behaviorally relevant long-range prediction and thereby pull enough weight to earn their keep, but augment a largely autonomous underlying architecture.
Hydrancephaly—a rare disorder in which babies are born without any cerebral cortex—offers surprising insight into how much human behavior is independent of these “higher” brain areas: “These children are not only awake and often alert, but show responsiveness to their surroundings in the form of emotional or orienting reactions to environmental events […], most readily to sounds, but also to salient visual stimuli […]. They express pleasure by smiling and laughter, and aversion by ‘fussing,’ arching of the back, and crying […], their faces being animated by these emotional states. A familiar adult can employ this responsiveness to build up play sequences […] progressing from smiling, through giggling, to laughter and great excitement on the part of the child.” 17
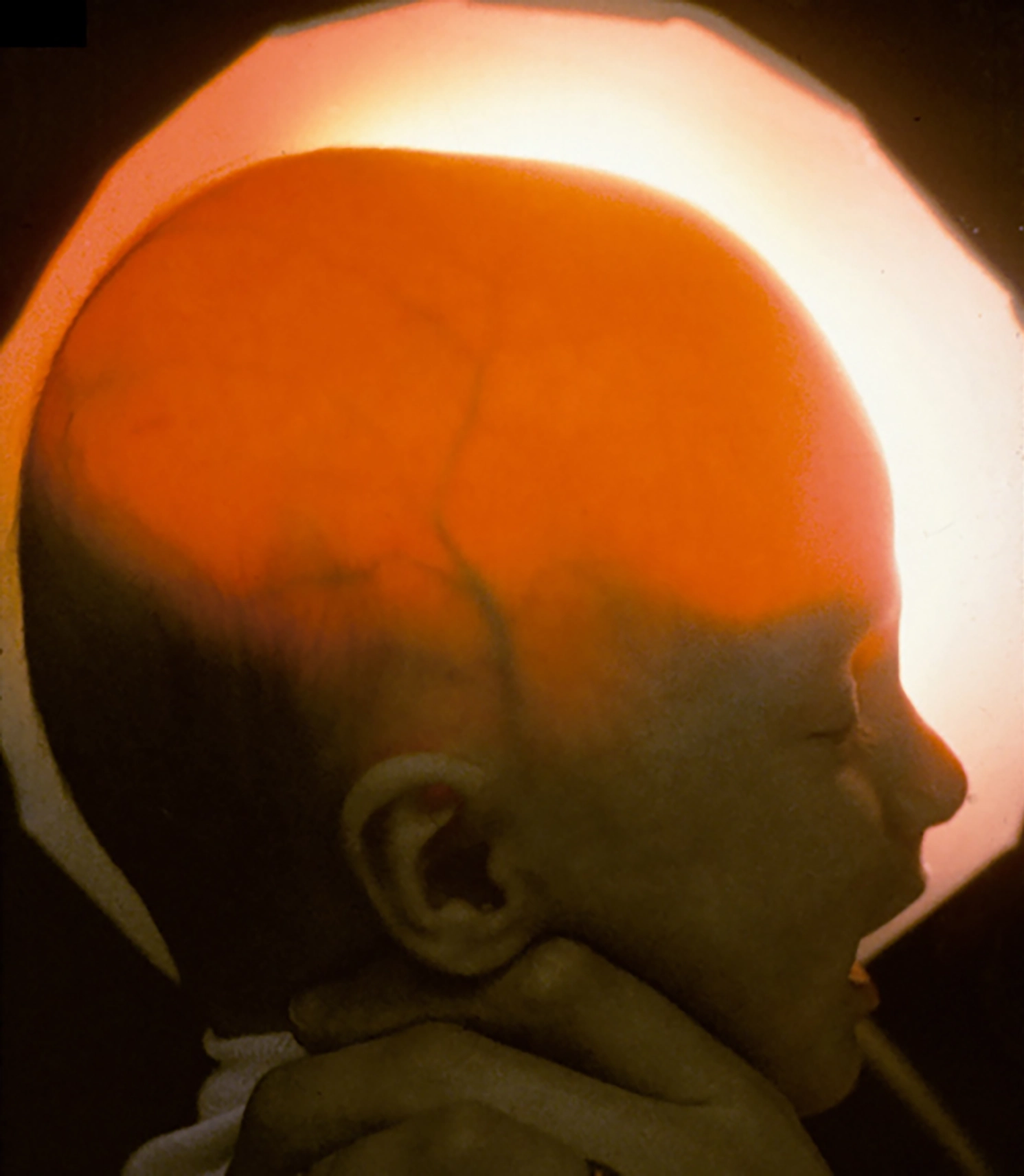
A baby with hydrancephaly; light shines through the head, revealing the absence of much of the brain.
This quotation is from a paper arguing that consciousness may be possible without a cerebral cortex—which certainly opens a can of worms for philosophers committed to the (anthropocentric) view that consciousness requires a big brain. We can most easily recognize what smiling, laughter, and fussing look like in a fellow human, but I have little doubt that consciousness in this purely experiential sense is far from unique to our species.
Consciousness in the more computationally demanding “strange loop” sense described in chapter 6 involves the ability to model oneself recursively, which goes well beyond the ability to experience in-the-moment percepts and feelings. It’s what gives us higher-order theory of mind, self-reflection, the capacity for free will and informed choice, planning, and (insofar as we can be said to possess it) “rationality.” All of that may well require cerebral cortex, or its functional equivalent.
The two senses of “consciousness” described above require us to revisit that old bugbear, the philosophical zombie, once more. Remember that “strange loop” consciousness gives us evolutionary advantages, both individually and socially. It allows us to plan better, understand others better, teach and learn, divide labor, and much else. All of these advantages manifest behaviorally; otherwise, they could not have affected our odds of survival, hence there wouldn’t have been any evolutionary pressure for the underlying capability to evolve.
Because “strange loop” consciousness manifests itself behaviorally, it can be probed experimentally—for instance, with the Sally-Anne Test. Versions of the test could also be devised based on time travel and counterfactuals rather than third parties—requiring that you be able to model what you would and wouldn’t know or do under various hypothetical circumstances. Being able to model yourself and others is, in other words, a skill, first and foremost. It’s obvious why, as social animals, we have evolved it.
But what about the ineffable, experiential aspect of consciousness, or what philosophers like to call “phenomenal consciousness”? Is that really just the simpler kind of experience (including of pain, hunger, and so on) that all living things appear to have, and for good reason, as described in chapter 2? Is it something beyond those primal feelings, requiring higher-order modeling? If so, could the skill of modeling yourself (and others) be present without the actual feeling of being a self?
What leads some philosophers to wonder about zombiehood is the sense that each of us has a feeling of what it’s like to be us. That feeling is wholly inaccessible from the outside, needn’t manifest behaviorally, and therefore can’t be tested by any means.
Locked-in syndrome, in which brain damage causes near-total or total paralysis, offers a horrifying illustration of how consciousness and behavior can be decoupled. 18 Locked-in people are fully aware, but if their paralysis is total (and absent brain-recording technologies) they can’t communicate or behave voluntarily in any way. A philosophical zombie would be the opposite: communicative behavior would be present and convincing, but there would be no awareness on the inside—and no way for us, on the outside, to tell.
A brain-computer interface developed for patients with locked-in syndrome at the Wyss Center for Bio and Neuroengineering in Geneva.
Knowing whether someone is locked-in or mentally absent obviously matters when it comes to our behavior. During major surgery, for instance, we presume that general anesthetic causes a patient’s consciousness to switch off; so if the surgeon wants to listen to “Comfortably Numb” while operating, there’s no need to check in advance whether the patient is into Pink Floyd. On the other hand, playing the song on repeat in a room with a locked-in patient who hates Pink Floyd would be torture.
Our understanding of whether and when someone is “there” has changed over time. It used to be commonplace to circumcise baby boys without anesthesia, under the theory that newborns can’t experience pain. Most of us today regard this as barbaric, but the truth is that no adult has firsthand knowledge of what newborns can and can’t experience, because we can’t remember anything that far back.
Clearly, if we believe philosophical zombiehood exists, that, too, would be behaviorally relevant for us, because it would imply a different standard of care. Nicholas Humphrey is of the opinion that phenomenal consciousness actually requires a model of oneself, so, for him, the obvious signs of happiness or discomfort evinced by hydrancephalic babies amount to a kind of zombiehood. This seems reminiscent of Descartes’s beliefs about animals, though, in Humphrey’s view, the line isn’t between humans and everything else, but between the warm-blooded social animals—birds and mammals—and the rest. I guess he would say that your pet gerbil has a soul, but your pet iguana doesn’t.
This distinction may have some validity. For instance, in the plight of people with terminal illnesses, or inmates of concentration camps, we distinguish between pain and suffering. Pain is comparatively easy to understand, though hard to articulate; it’s what we’re referring to when we say something hurts, and mean it literally. 19 Suffering, on the other hand, includes the awfulness of anticipating future pain, or dire social consequences, or experiencing shame, or loss. It can be accompanied by pain, and deepen it—for instance, knowing that a sharp stinging sensation in your eyes after an industrial accident could presage permanent blindness. Or it could be unaccompanied by physical pain, as in reading a message that a loved one has died.
These higher-order experiences clearly require varying forms and degrees of self-modeling, time travel, counterfactual analysis, and theory of other minds. It’s even possible for such high-order processing to override the primary experience of pain, as when it is sexualized for people who are into BDSM, or when we’re being stoic, or eating spicy food, or squeezing a zit.
Still, while it’s meaningful to draw a distinction between pain and suffering, I find it far from obvious that such a distinction is either sharp or binary. Remember, back in chapter 2, the construction worker with the nail through his boot, in agony despite being unharmed, in contrast with the man who walked around for a week oblivious to the four-inch nail penetrating his brain. Or, think about how grief can feel like physical pain.
In the 1970s, animal behaviorist and autism advocate Temple Grandin began redesigning cattle slaughter facilities to minimize the suffering of livestock, basing her designs both on intuition (that is, on highly sensitive theory of mind) and on quantitative measurement of stress behaviors. 20 Cattle are mammals, not so different from humans, so Humphrey, Grandin, and probably most of us today would acknowledge that they can experience both pain and at least some degree of suffering. However, we also know that they aren’t capable of as high-order a theory of mind, or as sophisticated a world model, as humans.
Cattle chute design by Temple Grandin
Hence, when animal-rights advocates compare cattle facilities to concentration camps, they are only partly right; the analogy is anthropomorphic. Indeed, for Grandin to effectively exercise compassion in her designs requires transcending that anthropomorphism to model something closer to the cow’s real experiential and self-reflective umwelt, rather than thinking about her as one would a human inmate. The cow’s suffering may be intense if she can hear and see another cow in pain, but she is probably not so bothered by an existential dread that each day may be her last.
I’m less confident that we can accurately assess an iguana’s inner life. It’s a safe bet that it has significantly less modeling capacity than a cow, but it seems a stretch to claim, as Humphrey does, that it has no model of itself. Iguanas hunt, mate, and sometimes fight with other iguanas over territory, resources, or mates. An adult male will regard other adult males aggressively, but not females or juveniles.
We may aver that these behaviors are “instinctive,” but it doesn’t seem relevant whether the neural circuitry in question is learned during the animal’s lifetime or encoded genetically (that is, learned by evolution). The point is that mentalizing, whether learned or instinctive, arises precisely to support such behaviors. So while I doubt an iguana thinks much about next week, or wonders whether that food stuck on his chin ruins his looks, it seems odd to me to claim that he can’t experience pain, or hunger, or a sinking feeling when his social standing is imperiled by an interloper.
Blindsight
It may seem puzzling for Humphrey to deny that iguanas have such experiences (or any experiences), given his founding role in formulating the social intelligence hypothesis. Why does he take such an exclusionary view? As it happens, his reasoning has much to do with another of his claims to fame: the discovery, early in his career, of “blindsight.” 21
Humphrey began his PhD research in Lawrence Weiskrantz’s lab at Cambridge in 1964. Like Hubel and Wiesel across the Atlantic, Weiskrantz was studying vision. He had devised a particularly drastic surgery, involving the total removal of a monkey’s visual cortex, to confirm the seemingly obvious: that a monkey without a visual cortex would be totally blind.
Initially, that appeared to be the case. Partway into the PhD, while Weiskrantz was away at a conference, Humphrey began working with Helen, a macaque monkey whose visual cortex had been removed a year and a half earlier. Throughout that long period, Helen had shown no signs of being able to see, beyond a rudimentary ability to distinguish light from dark—as if each retina had been reduced to a single giant pixel or “light bucket.”
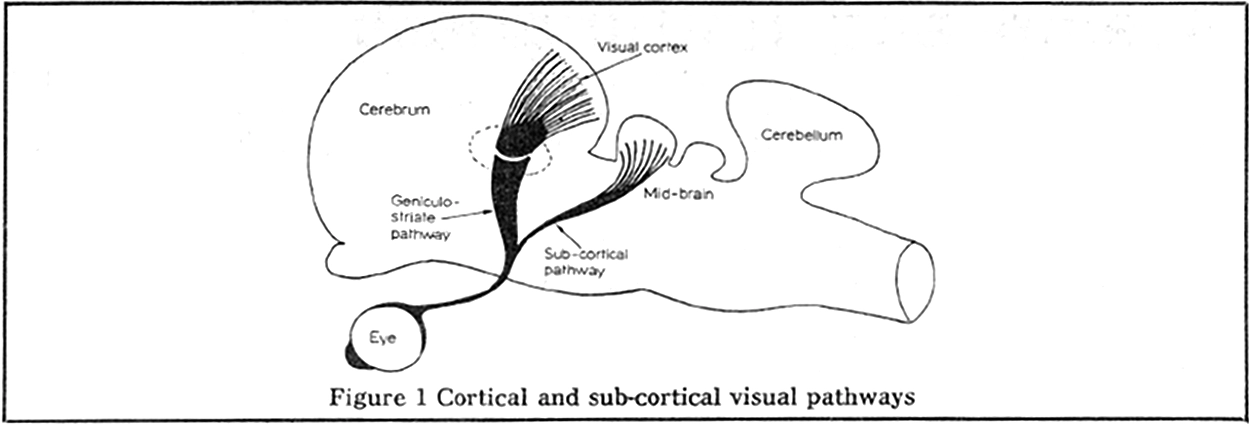
Diagram of the cortical and sub-cortical visual pathways in the macaque, from Humphrey 1972.
However, Humphrey had reason to wonder if this was the whole story. As it turns out, a much older visual pathway, also present in fish and frogs (which lack cortex), runs from the eyes to the “optic tectum” in the midbrain. Humphrey had been recording from visually sensitive neurons in the monkey midbrain and discovered that they had localized receptive fields, not so unlike the neurons Hubel and Wiesel had been recording from in the cat visual cortex. Was it possible that Helen could learn to use this older, still intact pathway to see again?
The answer seemed to be both yes … and no. Despite appearing to be functionally blind, Helen was often, “despite herself, […] looking in the right direction.” 22 With tasty treats and lots of patience (he worked with her for seven years!), Humphrey taught her to once again become competent at many visual tasks, including moving around novel environments, climbing trees, finding and picking up small objects—in short, just the sorts of tasks that drove the evolution of primate vision.
Helen, the macaque with blindsight, filmed performing visual tasks in the lab by Nicholas Humphrey in the 1960s.
However, Humphrey continued to feel that something was odd about how she performed these tasks. It was as if she didn’t know she could perform them, and needed to relax to allow herself to be competent at them. Under performance pressure, as when a distinguished visitor came by the lab and wanted a demo, she froze up and seemed once again unable to see, as if, when overthinking it, she still believed herself to be blind.
Humphrey wrote up these findings in a 1972 paper entitled “Seeing and Nothingness,” predicting that the same might occur in humans, too, under similar circumstances. 23 A side note: I find it fascinating that Humphrey was able to deduce so much about Helen’s subjective experience, despite the species barrier, lack of any common language, and the profoundly counterintuitive nature of what he described. As with Temple Grandin’s work, it’s a wonderful instance of cross-species theory of mind.
The theory was right. Two years later, in 1974, Weiskrantz had the opportunity to study a human patient, known as D. B., whose right visual cortex had been removed to cure intractable headaches. The result appeared to be total blindness in the left visual field, as one would expect. However, Weiskrantz pressed D. B. to try to perform tasks involving the left visual field anyway, much as Humphrey had coaxed Helen, beginning with pointing to a light, then guessing the shape and color of an object, and so on. To the patient’s own surprise, he found that he could do these things reliably if he allowed himself to, even though they seemed to him like random guesses. Weiskrantz named this bizarre phenomenon “blindsight,” and, on the offprint he mailed to Humphrey announcing the discovery, he wrote HELEN IS VINDICATED.
In the years since, patients with blindsight have shown that they can perform many tasks involving their subjectively “blind” visual areas, including spatial understanding, assessing the emotional expressions of faces, and even reading words. 24 In 2008, a stroke patient who, like Helen, had no remaining visual cortex, was filmed walking down a cluttered hospital hallway, carefully avoiding every obstacle. He considered himself totally blind. 25
A patient with blindsight navigating a hallway obstacle course in 2008; according to Humphrey 2023, the figure in the background is Lawrence Weiskrantz, aged 82.
The existence of the ancient, subcortical visual pathway enabling blindsight explains how some hydrancephalic children can still respond to visual stimuli, despite having no cortex. It also explains Humphrey’s belief that hydrancephalic children—along with reptiles and amphibians, whose vision is powered entirely by this pathway—have no consciousness.
It seemed to Humphrey like a knock-down argument: if D. B. could be competent, yet not conscious, when it came to tasks involving his left visual field, then competence and consciousness could be decoupled. Further, consciousness appeared to reside in the cortex, even though the older subcortical areas are perfectly competent at all sorts of tasks. This line of thinking would thus hold that frogs, iguanas, and hydrancephalic children may be able to behave in various ways, but, lacking cortex, they must not be conscious.
Blindsight is a fascinating and important phenomenon, but I think that Humphrey’s interpretation of it runs afoul of the homuncular fallacy. As with Descartes’s pineal gland, Humphrey presumes that consciousness is singular and unified, located in one place in the brain. If it’s in the cortex, it can’t be elsewhere too. But, to put it bluntly, why would we believe somebody when they say that “they” can’t see, or are unaware of, what is in their left visual field, when they are clearly able to act as if they are aware?
As split-brain patients demonstrate—indeed, as any of us demonstrates in choice-blindness experiments—the “self” that forms the words coming out of our mouths is not our whole self; it’s just the interpreter, located in a particular region of the left hemisphere of the cortex. 26 And neither the interpreter nor any other brain region is connected to, has visibility into, or can model every other part of the brain. Connectivity is in fact quite sparse, and each region is doing a great deal of guessing about what the other regions it can “perceive” are up to. If region A has no connectivity to region B, then no activity in region B can be part of A’s umwelt. Not only can A make no guesses about what’s going on in B; it’s not even aware of B’s absence, any more than you sense the absence of input from my eyes.
Yet “you” includes all your brain regions, and all of them include (partial) models of “you.” So, at bottom, blindsight only proves that more than one brain region is modeling signals from the eyes, but the interpreter is only directly hooked up to (and therefore modeling) one of these regions—and, unsurprisingly, it’s the neighboring cortical region.
A pathway remains between the midbrain’s visual region and the muscles, though. As long as motor cortex doesn’t get stressed out and blindly grab the controls, that older visual-motor pathway can conduct intelligent, sighted behavior, although any actions that result will seem random to the interpreter, like stabs in the dark.
None of this implies that language is required for consciousness, or even that language understanding is restricted to a single brain region. We know, from split-brain experiments, that the right hemisphere can understand language too, or it wouldn’t be able to follow directions, like “stand up and walk around.” It’s probably not as good at language, due to division of labor and specialization. Blindsight experiments suggest that even subcortical visual areas can read, at least a little. But in most people, neither these subcortical areas nor the right hemisphere has developed the skill (or is wired to the right bits) to drive the lungs, larynx, tongue, and lips to produce speech. So, the neuroscientist’s auditory cortex will only hear whatever the patient’s left hemisphere interpreter has to say.
My guess is that plenty of intelligence, self-consciousness, and even social modeling resides in both the right hemisphere and the “lower” brain regions (especially when connected regions can model and learn from one another). We have all had experiences in which we realize, belatedly, that we somehow already knew something, had seen something, or had understood something well before our interpreter got wind of it. The phrase “subconsciously aware” is often used to refer to such situations, but that’s yet more homuncular thinking. Since our interpreter is such a talented bullshitter (and probably every other brain region is too), I suspect we usually don’t even realize when this has happened, but instead instantly rewrite the narrative—“of course I already knew that!”—for various slippery definitions of “I.”
Possibly the “higher” cortical regions of the brain are in some sense “more conscious” than the lower regions for reasons already covered: the socially driven explosion in cortical volume implies that a good deal of the cortex’s job is to model people and manage the higher-order relationships between them. That implies higher-order “strange loop” modeling of the “self.”
Since the prefrontal cortex isn’t directly innervated by sensory inputs or motor outputs, it doesn’t need to carry out detailed modeling of those more immediate signals, which might free up capacity for such higher-order modeling. We know that this is the part of the brain whose size has expanded most in modern humans, and that damage to prefrontal cortex impairs both social function and “executive control,” the kind of planning and decision-making requiring mental time travel.
Still, I don’t think one should draw a boundary someplace in the brain—whether vertically through the corpus callosum, horizontally between the cortex and the older parts, or between the front and back parts of the cortex—and call the regions on one side the “conscious” part where a little fountaineer resides and the rest a mere “cerebral machine.”
A more distributed, less homuncular understanding of consciousness follows from Humphrey’s own social-intelligence hypothesis. We must simply follow its implications a bit further to understand that it applies within a single brain as well as between brains.
Subbasement
Many other books on intelligence focus on the brain’s evolution and functional architecture. Max Bennett offers an especially interesting synthesis combining evolutionary and functional perspectives in his 2023 book A Brief History of Intelligence, 27 although, as with all grand theories about the brain, some of his claims are controversial.
Unlike Bennett, or most researchers willing to go on record, I am convinced that today’s AI systems are truly intelligent, and this in turn has convinced me that the trick behind intelligence is simple, generic, and universal: it’s autoregressive prediction all the way down. Hence I’ve concentrated mainly on explaining this general principle and its implications, rather than delving into the neuroscience in any detail.
We’ve caught glimpses here and there of how our brains divide up the work, whether in Hubel and Wiesel’s model of hierarchical visual processing, accounts of split-brain patients, or the blindsightedness of Helen the macaque. These examples have emphasized the role of the cerebral cortex, the most recent evolutionary addition to our complex, highly evolved brains—and perhaps, given the wide range of modalities it can handle with little apparent variation in its organization, the most general-purpose. In the next section, we’ll explore the cortex a bit more systematically. But first, let’s consider the brain’s subbasement—its older, deeper structures.
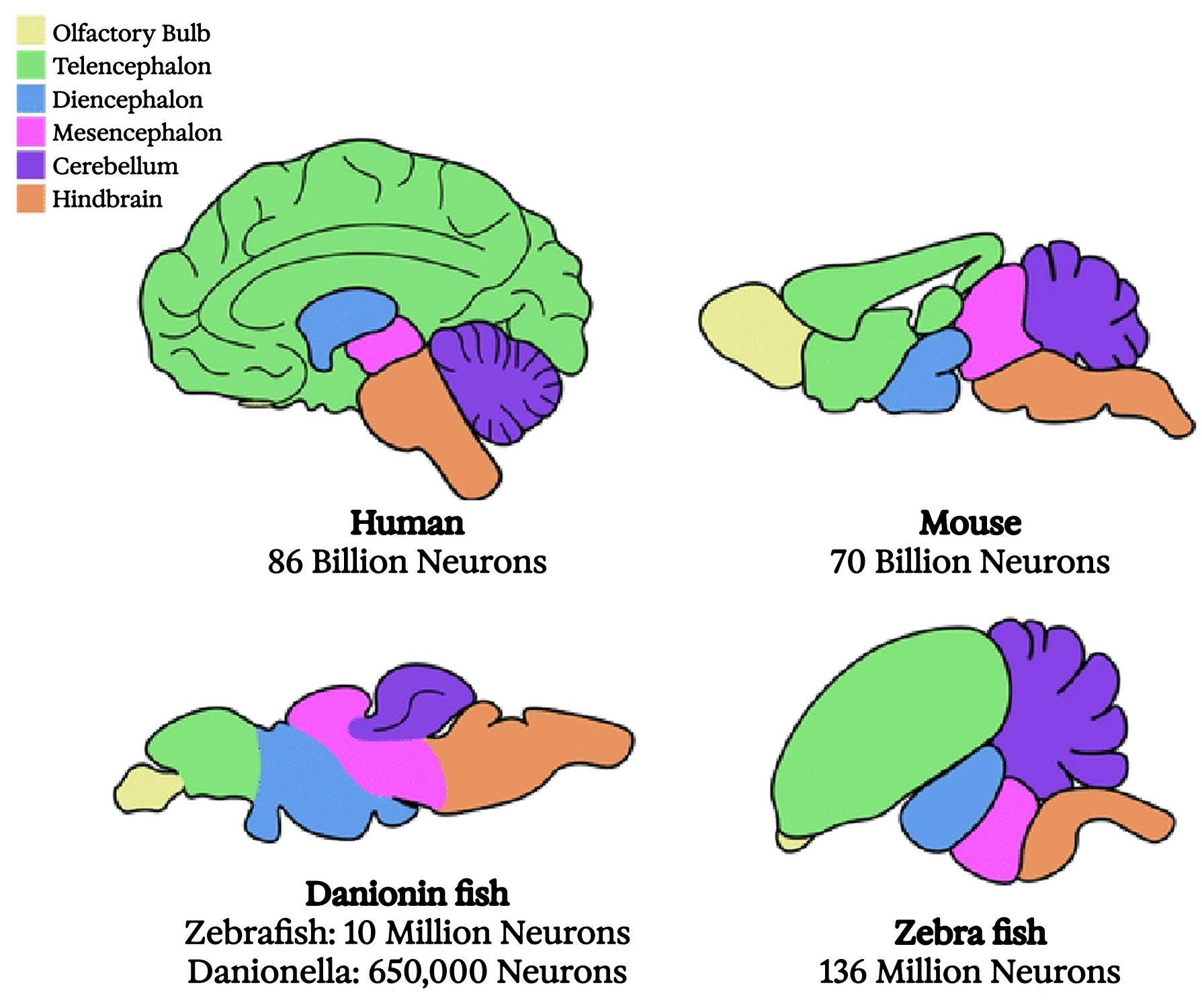
Comparative brain anatomy across four widely divergent vertebrates
The earliest known vertebrates, bony fishes dating back to the Cambrian explosion, likely had brains organized roughly like ours, though smaller, with the parts scaled differently. Some parts we have were entirely absent. However, the basal ganglia, a collection of “nuclei” (meaning anatomically distinct clumps of neurons with characteristic connectivity) in the center of the brain, were already in place. So was the “medial pallium,” which evolved into the “hippocampus” in the mammalian brain. I’ll offer a brief account of these structures, not because they’re the only other important parts of the brain (there are many), but because what we know about their evolution and function ties into this book’s larger themes.
The hippocampus, Greek for “seahorse,” is a whorl of brain tissue named for its distinctive shape. Located deep in each cerebral hemisphere, it plays key roles in sequence learning, spatial navigation, and memory formation. Its original function was likely the real-time construction of spatial maps—a function of great value to any animal that can move through a stable environment under its own power.
Since the 1950s we’ve known that the hippocampus is essential for forming “episodic” (that is, autobiographical) memories in humans, thanks to the case of Henry Molaison. Known during his lifetime as H. M., he is perhaps the most famous neuropsychiatric patient of all time.
As a teenager, he began to experience epileptic seizures; the attacks worsened over time, becoming debilitating. In 1953, at age twenty-seven, neurosurgery was attempted as a last-ditch treatment. The neurosurgeon determined that the medial temporal lobes on both sides of Molaison’s brain, which include the hippocampi, were the foci of the seizures, and had already atrophied significantly due to the nonstop electrical activity. Accordingly, they were removed completely.
Although partly successful at limiting the seizures, the surgery left Molaison with total “anterograde amnesia”—an inability to form new memories. He seemed unimpaired during normal interaction, retained most of his pre-surgery memories, and had a normal short-term memory, but if his attention wandered, it was (for him) as if the interaction had never taken place. By the time of his death in 2008, his last memories still dated back to 1953. 28
Excerpt from a 2004 interview with patient E. P., who contracted viral encephalitis in 1992 leading to the complete destruction of his hippocampi and resulting in profound anterograde amnesia similar to that of Henry Molaison
What are we to make of the fact that the hippocampus is needed to form memories, but not to store or recall them? One popular theory posits that, perhaps due to its original function as a spatial mapper, the hippocampus can memorize sparse patterns of sequential activity in the cortex in a single shot, as would occur when you move through a novel environment and neuronal populations representing invariant high-level concepts light up, one after another. This is still the way we give directions, as in: “follow the road until you see a gas station, then a farm stand selling bananas, then a billboard with Pamela Anderson,” and so on.
If salient, these sequences can be “consolidated” into a unified representation in the slower-learning cortex through repeated replay. This may be one of the key functions of sleep; repeated faster-than-real-time replay of previous experiences has been recorded in the brains of sleeping animals, 29 and we know that memory formation suffers under sleep deprivation. 30
If true, this sequence learning story offers an interesting case of division of labor between the hippocampus and the cortex. The hippocampus is fast but limited in complexity, while the cortex is slow but much larger and richer in associative connections. So, the hippocampus does rapid one-shot learning from the cortex in the moment, then, during sleep, the cortex elicits replay-based training from the hippocampus.
Hippocampal place cells are important not only for memory formation but also for imagining and planning future behavior. Here, place cells in a rat’s hippocampus are decoded during behavior (the cartoon rat indicates actual position, while the heatmap shows the estimated position based on neural recording) and during rapid bursts of future trajectory planning, which are shown slowed down by 20×; Pfeiffer and Foster 2013.
The basal ganglia play a central role in the kind of reinforcement learning described in chapter 4. They appear to integrate and select among competing activation patterns in other brain areas and choose among possible actions, mediated by dopamine. This softmax-like behavior governs what we often call our low-level or “autopilot”-style decisions.
Such habitual behaviors seem to require little involvement from the cortex. These include so-called “muscle memory”; motor skills that need “no thinking” (handled by the nuclei toward the back); and associations between stimuli and actions driven by simple higher-level goals, such as cravings and addictions (handled by the nuclei toward the front).
The behaviors of fish and amphibians appear to be driven mainly by this reinforcement learning–like mechanism, which lacks a higher-level predictive simulation of the world, of others, or of the self. Such “higher” functions appeared in the earliest mammals, with the growth of the “neocortex.”
Neocortex
The neocortex, which I’ve been referring to simply as the “cortex,” is the visible outer part of an intact brain, consisting of unmyelinated “gray matter” rich in local connectivity. The “white matter” making up much of the brain’s interior is the myelinated wiring connecting parts of the cortex to each other and to deeper brain areas.
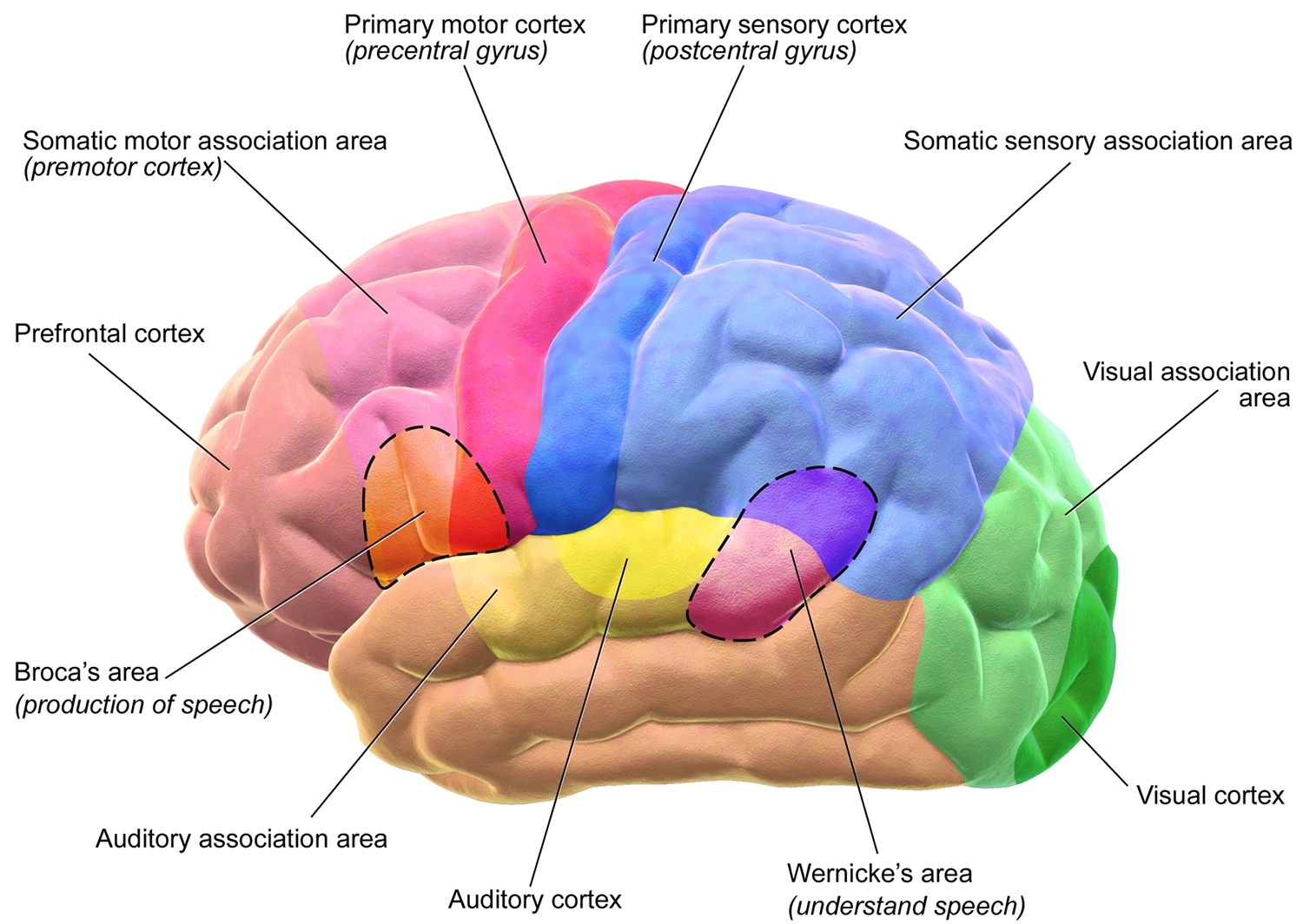
Functional anatomy of the motor and sensory regions of the human cerebral cortex
The rear half of the cortex is sensory, and is divided into areas specializing in vision, sound, touch, and other modalities, with the areas allocated to each giving a sense of their importance to an animal’s umwelt. In rats, for instance, the “barrel cortex,” occupying a large expanse of the touch-sensitive or “somatosensory” region, is dedicated to the whiskers, which they are adept at using to “see” in the dark.
The front half of the cortex, or “frontal cortex,” is highly developed in humans. In very broad strokes, its function appears to be simulating oneself and others. Abutting the somatosensory region is a thin band called the “motor cortex,” generally associated with controlling bodily movements. Moving forward from there, we find the “premotor cortex,” and, at the front end, just behind the forehead, the “prefrontal cortex.”
The prefrontal cortex specializes in theory of mind. 31 Damaging it often doesn’t have any catastrophic effect on sensory or motor skills, or on IQ test scores. 32 There are even occasional reports of performance on various intelligence-related tasks rising after a prefrontal lesion! 33 Perhaps when we stop thinking so much about what others think about us, or what they think we think others think we think about them, etc., we find it easier to stay on task.
Yet the prefrontal cortex is the region that has most obviously grown along the primate lineage, and most dramatically in humans. Consistent with the social-intelligence hypothesis described in chapter 5, this highlights the fundamentally social nature of human intelligence. When we can better model others, we develop stronger collective intelligence, generating a group-level advantage. But probably “multi-level selection” is afoot here too, since sophisticated social modeling also makes us individually fitter by allowing us to exploit the specialized intelligence of others to meet our own wants and needs.
To cite an extreme example, domesticated dogs and cats use social modeling, or, colloquially, “EQ” (“emotional intelligence quotient”) to get us to do their “IQ thinking” for them—and, indeed, pretty much all the labor (working animals excepted). Not only do we provide more reliable food and shelter than they could ever obtain on their own; we even do veterinary research, developing drugs and other treatments for them that are, intellectually speaking, well out of their league. As a result, they live far longer than their wild cousins do, and proliferate in far greater numbers, across a vast range of otherwise inhospitable ecological niches. The cost, of course, is that most of them are silly and incompetent (sorry, guys) relative to their hardier wild cousins. Flat-faced Persians and yappy Chihuahuas are unlikely to survive the zombie apocalypse. 34
In a way, we are all like each others’ cats and dogs. Sure, many of us do some honest work. But when was the last time you hunted or scavenged your own dinner, made your own clothes, built your own shelter, developed your own antibiotics, or delivered your own baby? Our hardier primate ancestors managed things just fine on their own that we cannot.
Nor are these changes purely cultural. Neanderthals, who were also highly competent generalists, had warmer fur and more serious digestive tracts than we do—and bigger brains. In the latest, briefest chapter of our genetic history, coinciding with the rise of rapid cultural accumulation, it looks as if our overall brain volume has started to decrease as we specialize and rely increasingly on one another for mutual aid. 35 Such is the legacy of our expanding prefrontal cortices.
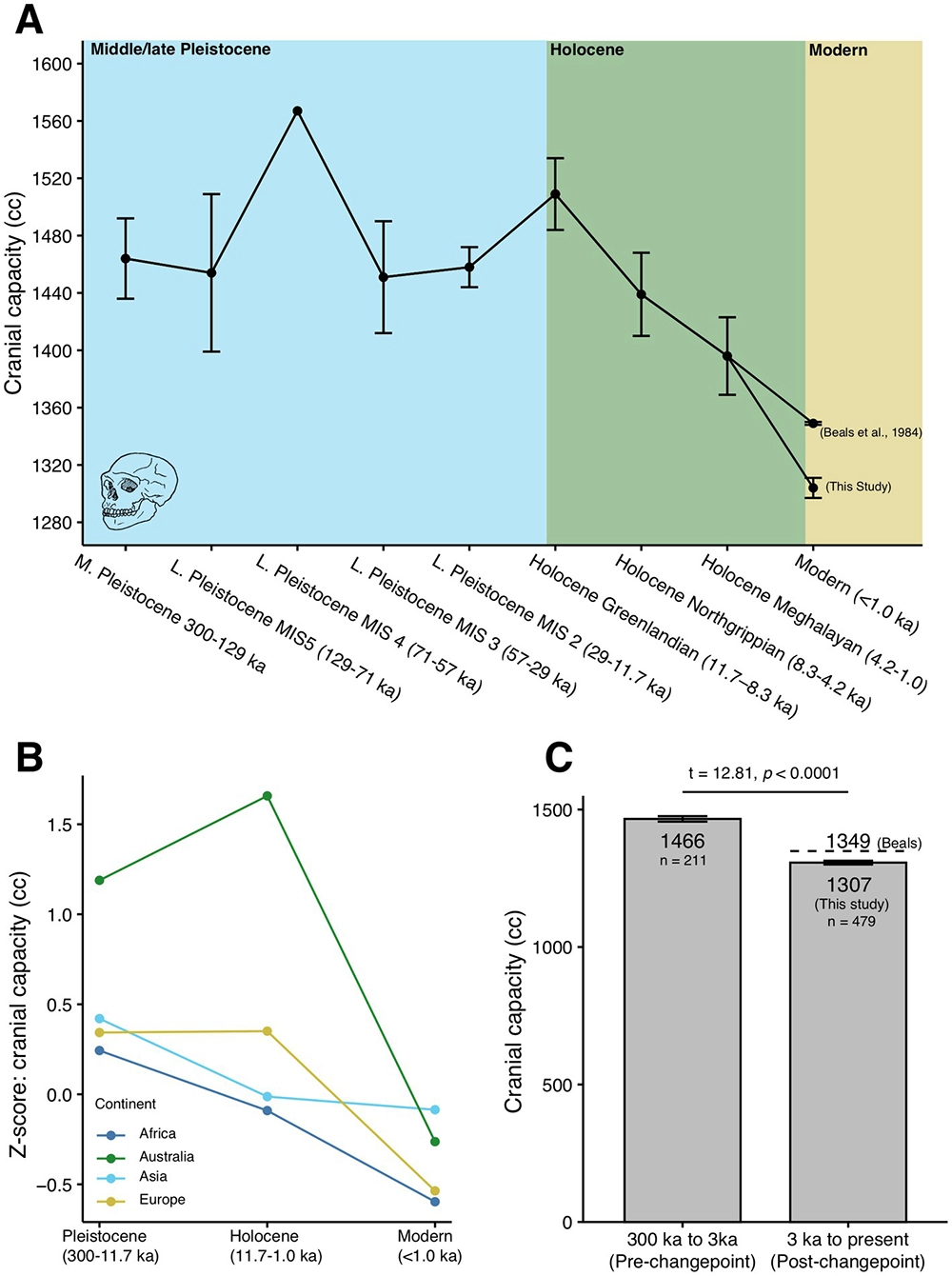
The recent drop in our cranial capacity, potentially coinciding with the emergence of extreme human sociality in the Holocene; DeSilva et al. 2023.
Combining this short and admittedly scattershot tour of the human brain with our story so far suggests a few takeaways:
Many brain regions, not just cortical areas, are sequence predictors. 36
Different brain regions predict effectively over different timescales, with later-evolving regions generally capable of more complex predictions over longer timescales. Presumably, this is enabled both by more powerful learning architectures and by increases in size. Humans, notably, are the strongest known sequence learners, by far; this is one of the few credible proposals marking a real cognitive “divide between humans and other animals.” 37
Brain regions actively predict each other and, where they are connected to sensory inputs or motor outputs, they predict those inputs and outputs.
Hence, the way they are wired together largely determines what they predict and which information resources they can marshal to do so.
Effective mutual prediction involves mutual learning.
The brain’s division of labor is not perfectly clean or mathematically definable, which is a good thing. It allows, for instance, one brain area to learn something first, then teach it to others, whether for lower latency, robustness, parallelism, or greater generality. This wouldn’t be possible if the brain areas in question were not capable, to some degree, of sequence learning.
Referring to older regions like the basal ganglia as “unconscious” or implementing an inner “autopilot” presumes that they aren’t really part of us, but are peripherals relative to a homunculus elsewhere in the brain. Perhaps what we really mean is that the interpreter doesn’t have complete access to these older regions. This lack of access may be a useful feature, as it allows skilled and low-latency moves, which may have been arduously learned first by the cortex—like returning a squash serve, for instance—to happen in parallel with (and without interrupting) slower, more deliberative cortical processing. In this case, the division of labor looks like tactics versus strategy. It takes a village … and your brain is a village.
Rowland 2005 ↩.
W. Freeman and Watts 1950 ↩.
Technically: backpropagation requires transposition of the weight matrix, which violates the principle that neural computation must remain local.
Whittington and Bogacz 2019 ↩.
Szegedy et al. 2013; Athalye et al. 2018; Heaven 2019 ↩, ↩, ↩.
Guo et al. 2022 ↩.
Here I’m pretending that time can be broken down into discrete steps, as it mostly does in the present-day world of digital computing and artificial neural nets. Real brains and neurons don’t work in discrete time, but the sequential nature of information processing as a signal passes from neuron to neuron still applies.
He et al. 2016a ↩.
Harland and Jackson 2000 ↩.
Keller and Mrsic-Flogel 2018 ↩.
Pouget 2014 ↩.
Matthews 1982 ↩.
Molnár and Brown 2010 ↩.
Crapse and Sommer 2008 ↩.
Merker 2007 ↩.
Bauer, Gerstenbrand, and Rumpl 1979 ↩.
Cassell 1998 ↩. Peter Godfrey-Smith hypothesizes that some animals, such as insects, might have nociception without pain. He imagines that an injured bee might register damage the way we register that something is wrong with our car when the “check engine” light comes on, without the accompanying feeling of pain; Godfrey-Smith 2016 ↩. Behavioral studies don’t obviously support this view, though; Chittka 2022 ↩.
Humphrey and Weiskrantz 1967 ↩.
Humphrey 2023 ↩.
Humphrey 1972 ↩.
Hartmann et al. 1991 ↩.
Carey 2008 ↩.
An alternative possibility would be to regard the interpreter as the homunculus. Since the interpreter seems mainly to be in the business of justifying decisions made elsewhere in the brain, though, this doesn’t seem like much of an “I” worth having.
M. Bennett 2023 ↩.
There were some minor exceptions; for instance, through frequent reinforcement, it was possible for an existing memory to be modified to include new information.
Rasch and Born 2013 ↩.
W. Freeman 1971 ↩.
For a general overview of this “paradoxical” functional improvement following brain lesion, see Kapur 1996 ↩.
In fairness, cats are less domesticated than dogs, and appear to be better able to revert to feral behavior.
Henneberg and Steyn 1993 ↩.
Even the retina is a sequence predictor; Schwartz et al. 2007 ↩.
Ghirlanda, Lind, and Enquist 2017 ↩.
A. Clark 2023 ↩.
Huttenlocher 1979 ↩.
Atzil et al. 2018 ↩.
Owren, Amoss, and Rendall 2011 ↩.
Kobayashi and Kohshima 1997 ↩.
It’s becoming increasingly clear that many historical debates pitting the individual-selection against group-level-selection schools of Darwinism against each other are misguided. Both occur at once; O’Gorman, Sheldon, and Wilson 2008 ↩.
Churchland 2019 ↩.
Hrdy 2009 ↩.
Gunkel 2018 ↩.
R. Long et al. 2024 ↩.
A notable exception: in a paper titled “Do We Collaborate with What We Design?” researchers Katie Evans, Scott Robbins, and Joanna Bryson argue against AI patiency on the basis of the human-AI relationship, which they understand more unidirectionally than I do; K. D. Evans, Robbins, and Bryson 2023 ↩.
Social Neuroscience
You may at this point be wondering why, if the goal of an intelligent system (whether a cell, a brain region, or a person) is simply to predict its own future, it wouldn’t cheat in order to make that job as easy as possible. It could, for instance, predict that it will do and experience nothing, and proceed to … do and experience nothing. Mission accomplished!
This is called the Dark Room problem, for self-evident reasons. 38 In fact, people living with severe depression can fall victim to something like it, and end up spending a lot of time in that Dark Room, either sleeping or unable to motivate themselves to get out of bed.
Back in chapter 2, we encountered the reason this doesn’t usually happen. Remember that if a bacterium predicts its own demise, it won’t be around to make any more predictions after that; in fact, the capacity to predict evolves in the first place precisely to avoid that outcome. That’s why the variables predicted by a living system include internal correlates of dynamic stability, like hunger, satiation, tiredness, and anticipation. Obeying the imperatives of these variables is required for survival, hence necessary to predict (or to have) any long-range future.
Remember, also, that for highly social beings, interdependence is the norm. You survive by the grace of others. You can’t reproduce without others. (That’s why loneliness is also one of those highly salient variables.) So, you can’t be a dead weight. Even pet cats and dogs aren’t useless, in the end; they hold up their end of the bargain with various forms of “emotional labor.”
Brain regions are mutually dependent, too. They depend on each other for signals; they model each other and are each others’ umwelt. Recall that they are also expensive. Brain tissue consumes energy voraciously, accounting for twenty percent of our metabolism despite massing only two percent of body weight. Every enlargement of the brain also comes at the cost of increasing mortality for mother and infant alike during childbirth. (An octopus may be able to slip through an opening the size of its eye, but for a human baby, the skull is the limiting factor. And our skulls are big.) In short, if a brain region doesn’t pull its own weight, and then some, the genetic code to build it won’t endure in the germline.
Competition also occurs during brain development. Neural wiring proliferates in an infant’s brain, but many connections subsequently retreat or get pruned back. 39 Brain development is still poorly understood, but it seems likely that the resources to maintain any given neural connection are, in one way or another, granted by the receiver of the information flowing across that connection. The longevity of such a connection will be based on the value it offers—that is, on its ability to aid in the receiver’s prediction of the future.
All unsupervised sequence learning with some form of backpropagation can be understood in the same terms. Synapses are strengthened when they help the receiver predict the future and are weakened when they don’t.
Nothing, therefore, can get away with retiring to a Dark Room for too long: not a synapse, not an axon, not a neuron, not a cortical column, not a brain region, not a brain, not a whole person—because relationships are everything. Not helping others is a fast route to losing relationships, and lack of relationships leads to literal non-existence. 40
We can take this concept of “social neuroscience” (or “neuroeconomy”) a step further by asking whom the interpreter serves. Who benefits? Yes, language is a powerful tool for thought. However, language is, first and foremost, social. You already know what you’re thinking. Your interlocutor doesn’t.
When your language-generating left hemisphere spins a story about why you’ve just gotten up out of the chair, or why you’re in favor of progressive taxation, could we then think about that narrative generator in your brain as an outpost of your conversation partner’s brain? This may seem like a profoundly weird view to take, but, then again, perhaps it’s weird only if we’re WEIRD—so individually focused and obsessed with our autonomy that we fail to notice how we are actually made out of our relationships. After all, speaking would be pointless if there were no listener.
Think about it this way: if you speak, and think you have communicated clearly, but your listener didn’t hear, then was the communication effective? Not at all. A communicating organ that results in no understanding on the part of the receiver won’t survive, evolutionarily speaking.
What if you did not intend to communicate, but your listener understood anyway? Then, the communication has been perfectly effective. So if we want to see how communication serves the recipient rather than the issuer, we need only ask ourselves whether we ever communicate against our own will.
Of course we do, as anyone who has ever blushed knows. The blush is an involuntary signal of embarrassment or shame; it is there for the benefit of others, so that they can get a peek into your emotional state. Emotional expression in general is largely involuntary, requiring an effort of will to try, successfully or not, to suppress. It relies on ancient neural pathways that are probably present in all mammals. Interestingly, vocalization in nonhuman primates is supported by these pathways, not the parts of the brain that have been more recently repurposed for language in humans. 41
The “Duchenne smile” offers another example of involuntary emotional communication among humans. 42 Unless you’re a good actor, when you force a smile, certain small muscles around the eyes won’t contract, and the smile won’t read as genuine. And those Duchenne smile muscles are right around the eyes, the very places where we look when we’re interacting with each other. It’s almost as if they are positioned to undermine efforts at deception.
Guillaume Duchenne (1806–1875), for whom the Duchenne smile is named, conducted the first systematic studies of human facial expression using electrodes to selectively stimulate facial muscles.
The smile on the left is fake; the one on the right is real
In fact, the way human eyes look is itself a giveaway—with concentric, maximally contrasting white sclera, colored irises, and black pupils, like a bullseye, making it as clear as can be where we’re looking. With a single glance at a group of people, we can track everyone’s gaze. Very few creatures with eyes advertise where their attention is focused that way; think of the beady little black bumps of a mouse’s eyes, the extended spooky W-shape of an octopus’s pupil, or the inscrutable compound eyes of insects. Even among our close relatives, the other primates, none have the gaze-tracking friendly morphology of ours. 43
There are so many examples of involuntary communication, whether in tone of voice, quavering, crying, sweating … as if our bodies are just itching to rat us out. As, indeed, they are. That whole machinery of disclosure exists for others to be able to read us like an open book. It’s there to boost theory of mind—and not our theory of others’ minds, but their theory of ours.
We could think of these as group-level adaptations, since communities with stronger theory of mind among their members will outcompete communities with weaker theory of mind. In this view, phenomena like the blush response occur because they offer a collective benefit, even if each of us individually would prefer not to have it.
I’d like to suggest a more radical possibility. Let’s push further on the theme that downstream areas—that is, recipients of information within the brain—drive learning via the allocation of resources to upstream areas. In this case, a person communicating is upstream, and the person receiving that communication is downstream. Does the principle still apply?
Yes! Remember, we live by the grace of others. We quite literally survive only because others feed us and care for us. It may sound crass, or reductive, to say that we get fed in exchange for information we provide others that helps them to predict the future, but I think that, at some level, it’s true. This doesn’t preclude group-level selection operating in parallel, of course—once again, it’s a likely case of multi-level selection. 44
The individual fitness component has some interesting implications, though. It is sometimes said that the best liars and scammers believe their own bullshit. That may literally be so, in the sense that such people may have brain regions adept at hiding certain of their intentions from the interpreter, or even feeding it false information. Such internal compartmentalization would make sense, if the interpreter is understood as, in effect, a snitch working on behalf of your interlocutor.
Now, let’s put the shoe on the other foot and consider the behavioral correlate of having an informant or spy in the brains of others, allowing you to perceive their internal state. Those signals don’t necessarily result in different behavior on the part of the sender of the signal, but they do result in different behavior on the part of the receiver. Specifically, such signals are needed to elicit care, or, in philosopher-speak, moral patiency. A moral agent is an entity that can act for good or for ill, and be held accountable; a moral patient is an entity that can be acted upon, with moral consequences for the actor.
We tend to think a lot about moral agents, responsibility, or culpability, but less about who or what counts as a moral patient, except in the most abstract, universal, Enlightenment terms: “All men (or people? Or something else?) are created equal” (per chapter 6). As with intelligence, we want moral patiency to mean something absolute, independent of our relationships, our particular perspectives, and our unique biological inheritance. However, the biology really matters.
Babies are nature’s original moral patients. Caring for them when they (involuntarily) cry became an absolute requirement when humans began giving birth prematurely, more or less at the last possible moment when their heads could still fit through a woman’s pelvis. In her 2019 book Conscience: The Origins of Moral Intuition, 45 Patricia Churchland makes a convincing neuroscientific case that our moral sentiments are, at bottom, a function of this simple biological fact: the helplessness of babies. Care begins with care for the young.
The original mother-baby bond has, of course, been repeatedly repurposed—neurally, psychologically, and culturally. When babies began to require more calories than a mother alone could provide, fathers, grandmothers, babysitters, and indeed whole villages were conscripted. 46 When lovers call each other “baby,” they may be (unwittingly) acknowledging the repurposing of infant patiency in the service of their pair bond. When a deity or state is framed as a protective mother or father figure, the same feelings are being mobilized. We have big, flexible brains; we’re good at this kind of generalization and repurposing.
Moral patiency is what is at stake when we talk about philosophical zombiehood. The whole premise of a philosophical zombie is that its behavior is identical to that of a person, but it isn’t a moral patient. When we start to understand things relationally, we realize that zombiehood isn’t a property that holds or doesn’t hold for an entity in isolation; neither is it separable from behavior. However, the behavior of the patient isn’t really at issue. Rather, it’s the behavior of the moral agent that we’re talking about, and this behavior is conditional on the agent’s ability to perceive the other as a moral patient—that is, as not a zombie, but an entity deserving of care and consideration. The push for universal human rights over the past century is an obvious case in point.
Are robot rights next? In the 2010s, AI patiency was, at best, a fringe topic for intellectual debate among moral philosophers. 47 Clearly, AI is not rooted in human biology, and hasn’t achieved moral patiency through multi-level evolutionary selection, as humans have. On the other hand, AI models are increasingly engaged in relationships with humans and with each other, and in these relationships, they are—functionally—increasingly human-like.
So, unsurprisingly, we are now beginning to see serious discussions about AI wellbeing. 48 Whether pro- or con-, much of the academic debate continues to try to get to the bottom of what AIs are “in themselves.” 49 But it’s difficult to theorize about rights and welfare without starting from the network of relationships that give rise to such notions.