Forking Paths
We’re finally ready to begin tackling the big questions posed in the introduction: the nature of general intelligence, the “hard problem” of consciousness, and the thorny issue of free will.
Let’s begin with free will, picking up where chapter 3 left off, with Nvidia’s DAVE-2 self-driving car. Our goal will be to understand what we mean when we call a model’s actions “willed.” Recall that DAVE-2 was nothing but a convolutional neural net whose input was a front-facing camera and whose output was a steering-wheel position. It was the purest and simplest approach to cybernetic self-driving. But is it capable of choice?
More concretely, what happens when such a “reflex-based” car arrives at a T-junction? A generic, pretrained model operating without any larger context can’t react sensibly. When we’re driving, these are the moments when we might make decisions based on higher-order goals. At other times, we’re on “autopilot” ourselves: the goal is merely to stay in our lane, follow the rules of the road, and avoid collisions with other vehicles, pedestrians, or animals. If we’ve established habitual routes, say to and from work, we might also make a lot of turns on autopilot. Those are instances when our inner “autopilot model” is not generic, but specific to our own lived experience.
DAVE-2, however, is only trained to perform the generic autopilot task. To gather the dataset, human drivers navigated a wide variety of routes on many different roads, and excluded from the training data the brief intervals when they were changing lanes or making turns. During live road tests, the human driver intervened to perform these maneuvers manually.
What would the car do if kept on autopilot at a T-junction? Presumably, it would choose the straightest way. At an exactly symmetric T-junction, one would hope that the model itself is just a tad asymmetric, causing it to break the tie somehow. A perfectly symmetrical model would be unable to break the tie, hence would be forced to generate the only non-tiebreaking output: steering straight ahead and running off the road. (Remember, DAVE-2 doesn’t control the gas or brakes.)
This unfortunate situation illustrates the old saying about the perfect being the enemy of the good. It also recalls “Buridan’s ass,” often framed as a philosophical paradox in discussions of free will. In the parable, the ass (or donkey, though the other definition applies too) is midway between two equally sized piles of hay, and, being unable to decide which to eat, starves to death. 1
Avoiding this “metastable” situation is easy enough. All it requires is a random variable, a bit of noise in the system, to break any near-ties.
In real life, though, Buridan’s ass only rears its poor muddled head infrequently, because few decisions we make are truly arbitrary, fully symmetric, or contextless. We don’t need to make random turns at T-junctions since, when we’re driving, we’re usually trying to get somewhere. More broadly, questions about choice, decisions, and free will only make sense when we consider an agent with its own history, acting in time and in context. That’s why we don’t generally count generic reflex actions as willed.
DeepMind’s AlphaGo contest with Lee Sedol took place in 2016, the same year Nvidia’s DAVE-2 self-driving-car paper was published. Go is exceedingly complex, with many more possible moves at any point in the game than chess—a vast and intricate garden of forking paths. For this reason, it had resisted the brute-force search or heuristic approaches that had produced grandmaster-level chess programs many years earlier, as well as naïve temporal difference reinforcement learning.
Comparison of Monte Carlo tree search in chess with the much larger number of possible moves in Go; Kohs 2017.
At bottom, DeepMind’s trick was the same as Nvidia’s: replacing hand-written code with a learned convolutional neural net. AlphaGo’s system design was considerably more complex, though, not only involving separate policy and value networks trained through reinforcement, but also incorporating traditional methods, like a randomized or “Monte Carlo” tree search through possible future moves. 2 In this sense, it was a hybrid between deep learning and GOFAI, though subsequent models—AlphaZero, 3 MuZero 4 —moved progressively toward a purer deep-learning approach, removing the remaining handcrafted heuristics while further improving performance. MuZero is general enough to play any game, not just Go.

Demis Hassabis as a young chess player
DeepMind has always aimed to solve artificial general intelligence through an agential approach. Gameplay runs deep in the organization’s culture. Its founder, Demis Hassabis, was a chess prodigy and an expert at many other games. (In 2024, he won the Nobel Prize, along with John Jumper and University of Washington researcher David Baker, for AlphaFold, which applies deep learning to solving the longstanding problem of predicting a protein’s structure from its amino-acid sequence.) Prior to DeepMind, Hassabis designed video games, and even founded his own gaming company. DeepMind’s first highly visible success, in 2013, involved combining reinforcement learning with convolutional neural nets to play Atari games. 5
Progression in the skill of DeepMind’s reinforcement learning agent playing the Atari game Breakout after one hundred, two hundred, four hundred, and six hundred training episodes; Silver et al. 2013.
The company’s bet that gaming could provide a controlled arena for developing general AI has certainly yielded impressive and important advances. Go commentators studying AlphaGo’s games lauded the system for “exhibiting creative and brilliant skills and contributing to the game’s progress.” 6
However, beyond the limitation Churchland pointed out 7 —its single-mindedness—AlphaGo was very different from the brain in another important way: it did not exist in time. It had no dynamics or internal state.
Children of Time
Like the self-driving-car model, neither AlphaGo nor its successors maintain any explicit memory of its prior “thoughts” or plans. 8 True, the system works out its next move by thinking as many steps ahead as possible, making it a stretch to call that extended evaluation a “reflex action.” However, once the move is made, it’s groundhog day—the next turn, it considers the board afresh.
AlphaGo makes no attempt to model its opponent’s psychology, nor does it learn on the fly; thus, it can’t individuate. Neither can it have any sense of a “self” acting in the world. All it models is that world itself and the possible paths through it, annotated (courtesy of the value network) with each path’s goodness or badness.
For a board game like Go or chess, none of these design choices are necessarily shortcomings. According to game theory (as formalized by none other than John von Neumann, 9 who had his fingers in many pies), the optimal next move depends neither on history, nor on the opponent, nor on any internal state, but only on the state of the board and the rules of the game—to which both players have full access. And the opponent should be presumed to always play optimally. To do otherwise would risk falling prey to a wily antagonist, who might lull an opponent by playing dumb, then take the gloves off at an opportune moment. 10
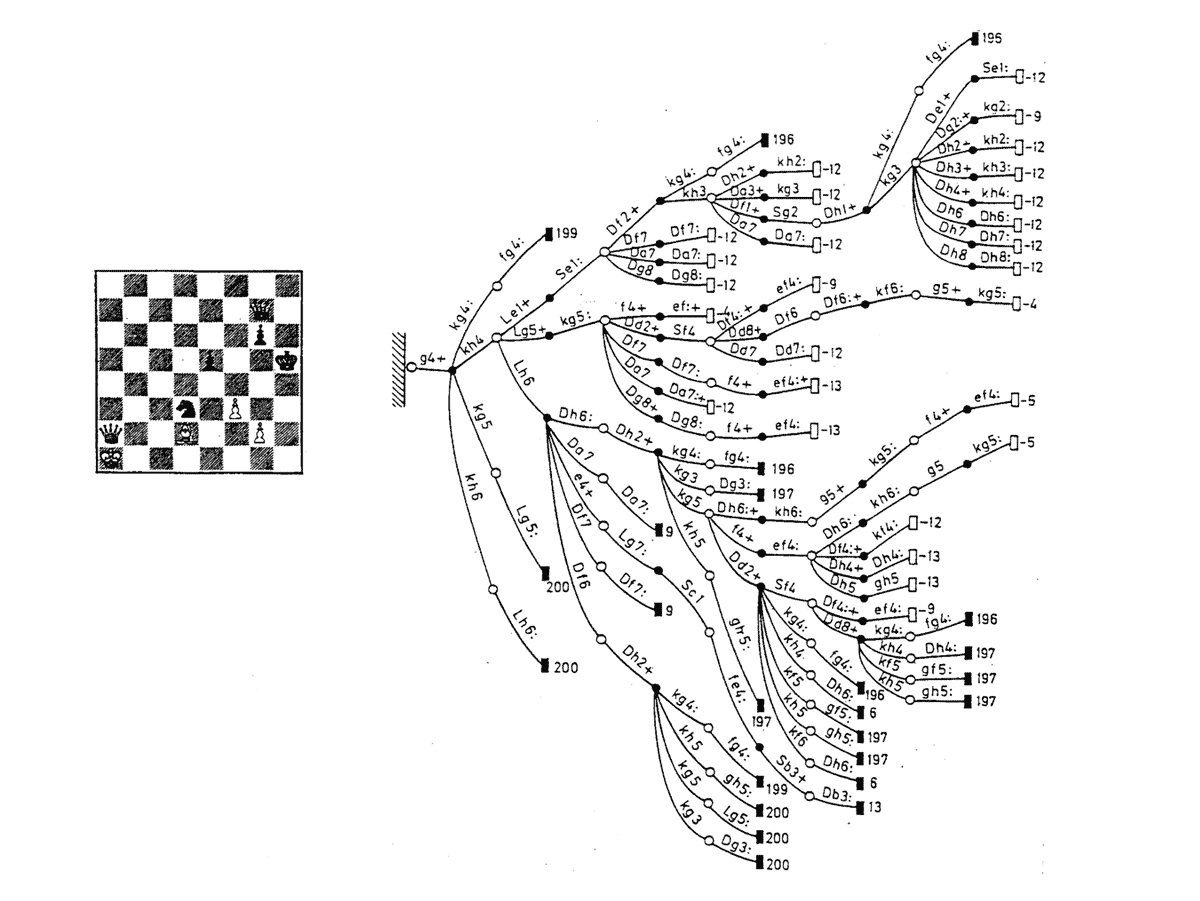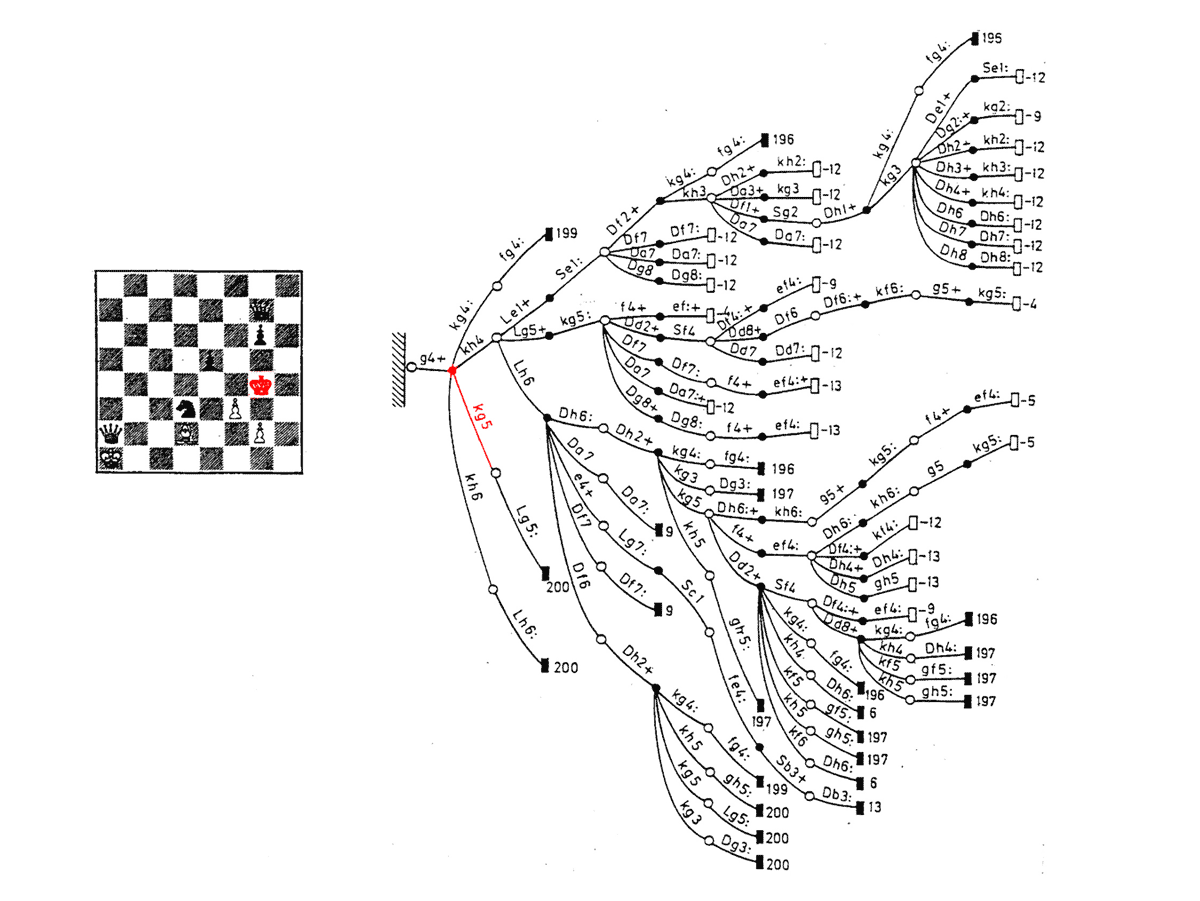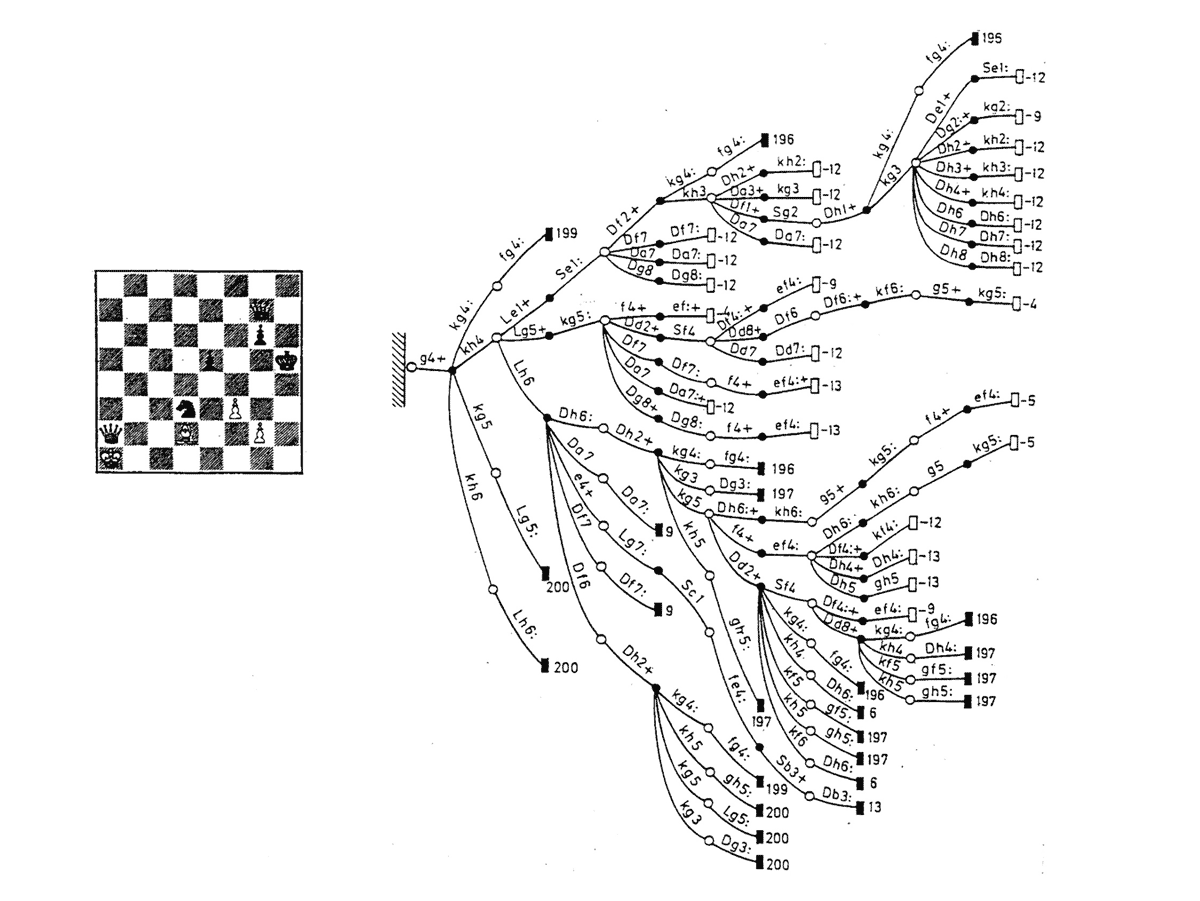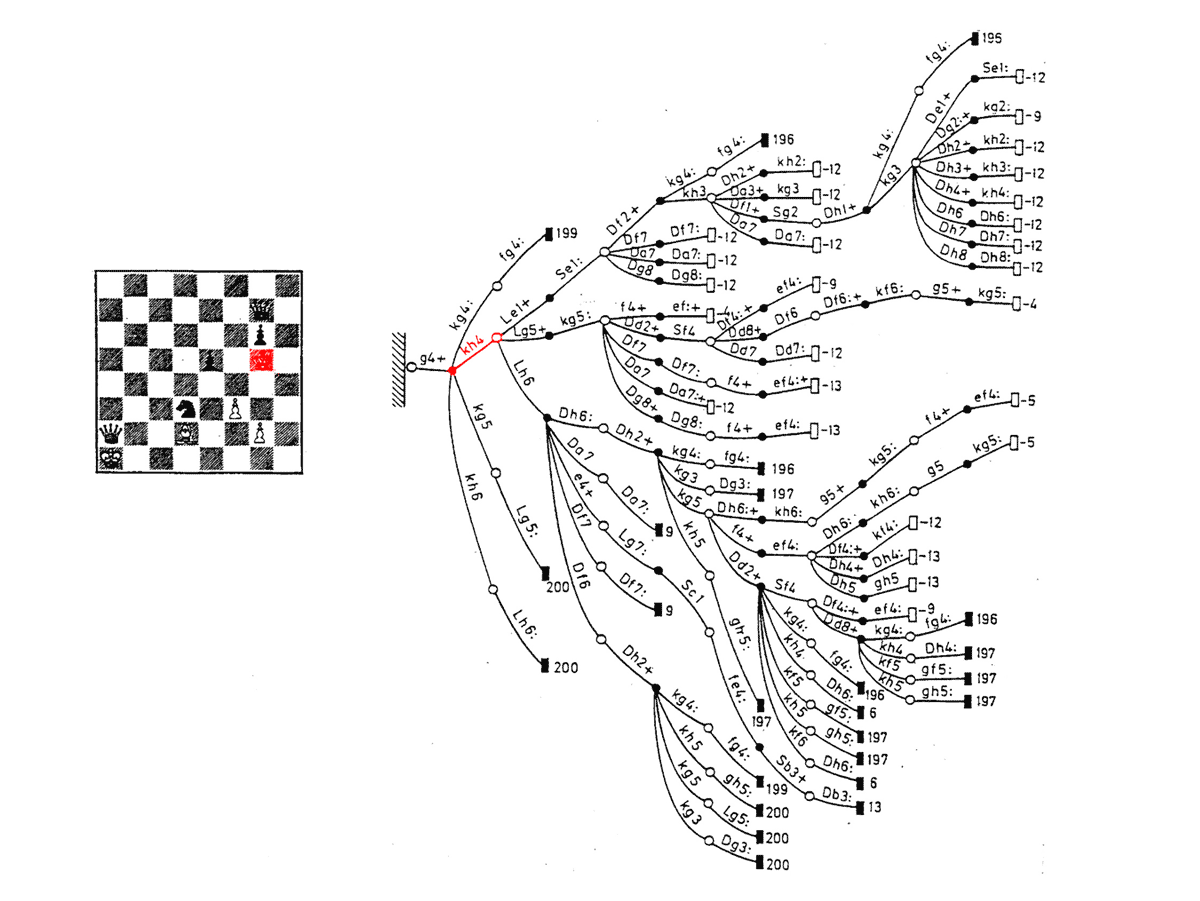
Decision tree of a chess study; Botvinnik 1970
You may wonder how AlphaGo could pull off a complex, multi-turn strategy—which it certainly can—without a self or memory. But given a clear view of the future’s forking paths at every turn, there’s no need to remember. All possible strategies are visible at all times, including the continuation of whatever strategy seemed best on the previous turn. If it’s still the best option after the opponent’s move, then so be it.
While elegant in its way, this approach can be wasteful, in that it often involves rediscovering the same strategy, turn after turn. Occasionally, catastrophic (and quite nonhuman) failures occur, since the model’s view of the future is not perfect, but, as with all Monte Carlo methods, stochastic. The model might make an unusual move due to glimpsing a brilliant but distant outcome down the line, but later fail to spot that same opportunity, even if it’s still in play, and foreclose on it with a wrong move. Out of sight, out of mind. 11
Movement and behavior of Portia
Portia spiders offer an illuminating contrast to this kind of “statelessness.” Although only a centimeter or less in size, they have been called “eight-legged cats” due to their unusually intelligent hunting behaviors. 12 They’re odd-looking, with a slow, choppy, robot-like gait—until they pounce. Unlike other spiders, Portia have a pair of large, forward-looking camera-type eyes with high resolution over a narrow field of view, well adapted to nuanced visual discrimination. Their favorite meal is other spiders, typically web-weavers, which may be twice their size. Portia can control the behavior of prey by spoofing web vibration signals; they learn which signals generate the desired response for each species through trial and error. In the lab, they can even learn how to fake out species they would never encounter in the wild.
Portia hunting
They plan their attacks slowly and carefully. With tiny brains and telephoto eyes that need to scan around to resolve the environment, it seems likely that they sacrifice much of the parallelism of animals with bigger brains (a topic we’ll return to in chapter 8). Once they have worked out a strategy, though, they can execute it virtuosically, leaping from twig to twig, abseiling down silk threads, and landing on their target from above, behind, or frontally, depending on what is safest under the circumstances (for oftentimes, the spider or insect they’re stalking is perfectly capable of eating them if the tables turn). Perhaps most impressively, Portia routinely reach their prey via indirect routes, including detours of up to an hour during which they may move away from their goal, breaking line of sight. 13 Impressive stuff!
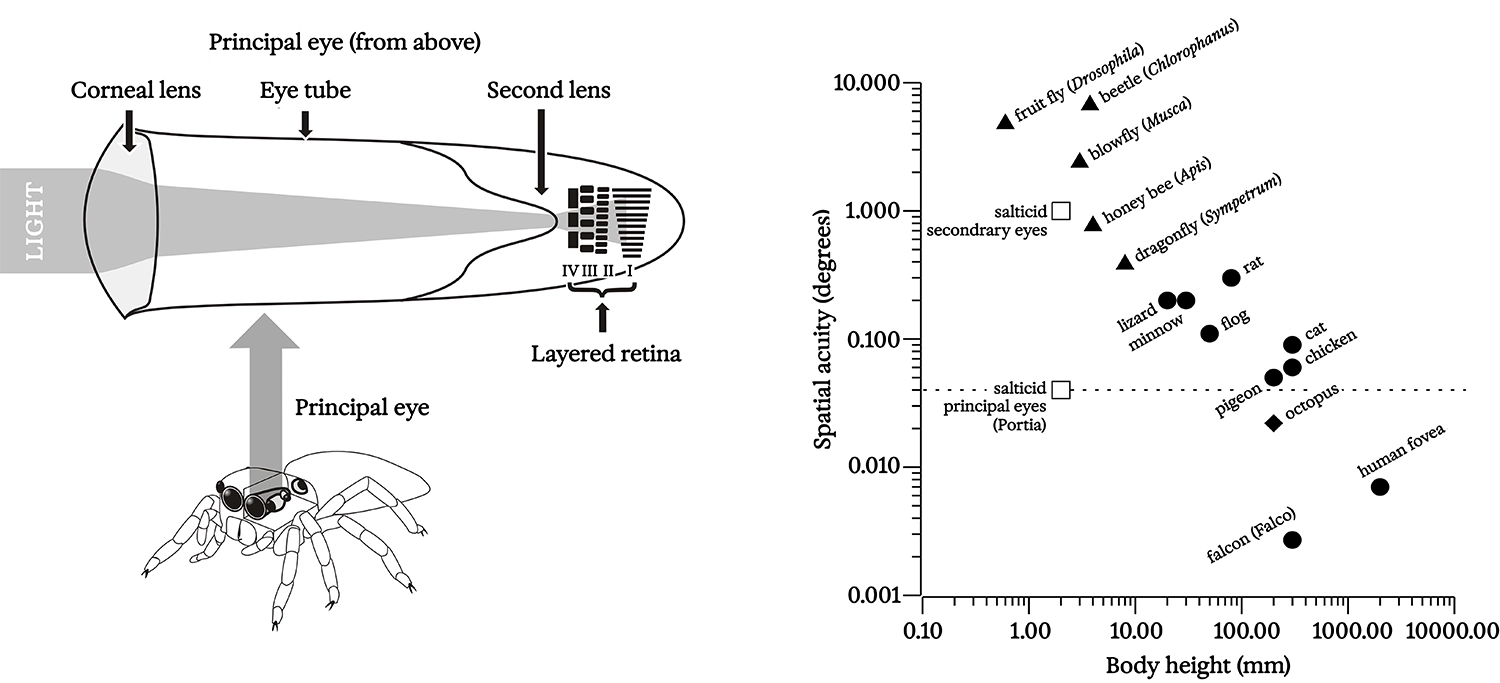
Structure and acuity of Portia’s principal eyes; Harland and Jackson 2000
I’ve offered this detailed account because it illustrates—and builds on—a number of Churchland’s observations:
- Portia isn’t playing any one game against any single opponent. Eventually it must eat if it is to survive, but it gets to choose with whom to engage, and when; it can always back off and try something else, or “soldier on.”
- The “game board” is not equally visible to all players. In its totality, it isn’t visible to any player. Portia must build up and incrementally update a mental model of the world in order to make decisions.
- This model must include not only the externally visible world, but the internal states of hunter and prey alike. Knowing its own state allows Portia to assess how hungry (or desperate) it is, how strong or weak it feels, whether hunting should take precedence over mating or resting, and so on.
- Modeling the mental state of its prey is also essential. Are they aroused? Have they realized I’m stalking them? If so, how well have I been localized? By timing its movement on a victim’s web to coincide with wind gusts, Portia may work to remain “vibrationally invisible”; or, by strumming the web, it may attempt to create the illusion of a trapped bug. The trickster must constantly assess: are my tricks working? Is this getting the reaction I want?
None of these tricks or capabilities are fundamentally out of reach for artificial neural nets; after all, each capability can be expressed in terms of functions, and neural nets are universal function approximators. Such capabilities will not, however, emerge on their own in a system like DAVE-2 or AlphaGo, due both to the constraints of the setup (autopilot or board game) and the structural limitations of a stateless classifier—even one that makes very clever decisions in the moment by evaluating many possible futures.
Sphexish
Given Portia’s alien lifestyle, physiology, umwelt, and brain limitations, it takes a leap of imagination to understand what it’s like to be a jumping spider, though it is fun (and maybe fruitful) to make the attempt. 14 My own sense is that Portia, cats, and humans on the hunt all share not only the ability to assess and plan an attack (or defend against one), but also to experience doubt, uncertainty, fear, and triumph. I expect that such feelings and experiences would emerge naturally from the evolution of predation, as described in chapter 3, just as hunger emerges naturally from the need to eat, as described in chapter 2.
But it seems so subjective to assert that another mind is experiencing something familiar from one’s own experience. How can one really know? We will soon explore how such claims can be assessed more rigorously, as predictive hypotheses.
For now, though, let’s stick to a more ethological perspective. Ethology concerns itself with animal behavior, studied using lab experiments and analyzed in the context of neurophysiology, function, and evolution. Through the ethological lens, we can simply ask the same questions about feelings and internal states we would ask about physical features, like the differently adapted beaks of Darwin’s finches. Why would that feeling arise and be preserved by evolution? How is it useful? What would happen if it were absent? This lets us fend off charges of anthropocentrism and defer “philosophical zombie” questions a little longer.
Free will, as a desire or imperative, seems easy enough to understand from an ethological perspective. No animal likes being trapped, humans included; that’s why imprisonment is a form of punishment, even if our bodily needs are met. Remember, the whole point of having a brain is to be able to act in the world, choosing among alternative futures in order to enhance our dynamic stability—that is, to stay alive, and to continue to be able to make choices in the future.
When others restrict our ability to make choices, it doesn’t feel good, just as gnawing hunger doesn’t feel good. The confinement of our possible futures renders us increasingly helpless, like a king being chased into a corner of the chess board in the endgame.
Go offers an even purer expression of this idea. Although the stones don’t actually move once laid down, their “liberties” are the number of adjacent unfilled board positions where they “could” hypothetically move, and stones only remain “alive” as long as they have at least one liberty. Death, then, whether in a game or in real life, represents the ultimate exhaustion of one’s ability to choose. It’s the pinched-off end of our Wiener sausage. 15
Mutual prediction is the key to sociality and cooperation, but, in a predatory context, as the early cyberneticists understood, prediction can also be adversarial. A predator will try to predict prey right into its stomach. Prey will try to predict the predator’s moves, and escape. Each is trying to preserve its own liberties, potentially at the cost of the opponent’s. Each will therefore try to predict the other’s predictions, ad infinitum.
A mouse trained to get from the entrance (left) to the exit (right) of a complex arena cluttered with visual occlusions outsmarts the robot chasing it using theory of mind; Lai et al. 2024.
As we’ll discuss in more detail in chapter 7, when mutual prediction is cooperative, it must also be imperfect, for cognitive cooperation involves a division of labor. That means each party brings cognitive resources or inputs to the table that the others lack. If one of the cooperating parties is always perfectly predictable to any of the others, that would imply that the predicted party doesn’t bring anything unique to the table, and is therefore a third wheel.
So, whether in a cooperative or adversarial context, being perfectly predictable is bad news. In a way, it’s even a form of imprisonment. Ted Chiang mordantly explores the psychological consequences in his short story “What’s Expected of Us.” 16 A device called the Predictor, consisting of nothing but a button and a light, appears on the market. The light always flashes green one second before the button is pressed. Millions of Predictors are sold, and, at first, people treat the device as a novelty, showing it to their friends and trying to fool it. When the implications sink in, though—that the Predictor can’t be fooled—free will is revealed to be an illusion, and people despair, eventually losing their will to live.
Real-life predictors aren’t single-button devices, of course, but other minds, like that of a cat pursuing its prey. And one way to become highly predictable is to run out of voluntary moves, like a cornered rat. But if one’s behavior looks deterministic from the outside—if it can be perfectly predicted by an observer—then, from an adversarial perspective, that is equivalent to being cornered.
When an agent becomes perfectly predictable to an observer, it makes the agent no longer register as agential to that observer. Cognitive scientist Douglas Hofstadter has described this property as “sphexishness,” by reference to the golden digger wasp Sphex ichneumoneus. 17 Hofstadter was drawing on observations by French entomologist Jean-Henri Fabre (1823–1915), who has been hailed as the founding father of ethology. 18 Although generally a keen and sympathetic observer of animal behavior, Fabre took a dim view of cognition in insects, claiming they exhibit a “machine-like obstinacy.” 19

Jean-Henri Fabre in 1880
He noticed that when the time comes for Sphex to lay eggs, she constructs a burrow and seeks out a cricket to nourish her future hatchlings. Instead of eating the cricket herself, she paralyzes it with a sting, brings it to the burrow entrance, goes in to see that all is well, then drags the cricket in, lays the eggs alongside the body, seals the burrow, and flies off. This elaborate performance certainly looks purposive—and it is.
A Sphex wasp brings a cricket to the entrance to her burrow, goes inside to check that all is well, and comes back out to drag the cricket inside
But then, Fabre noticed that if the cricket is moved by a few inches after the wasp has gone into the burrow to check that all is well, Sphex will re-emerge and move the cricket back into position, then go back into the burrow to check that all is well again! He described repeating this process dozens of times. By moving the paralyzed cricket every time the wasp went into the nest, Fabre was able to get her stuck in an endless loop. 20
One interpretation of this little parable: it’s easy to be fooled into thinking that a behavior is intelligent or agential when in fact it’s just a running program. That would beg the question, though: what else could it be? In some sense we are all “just running programs.”
A better take is to recognize, first, that Sphex’s nest preparation behavior is genetically determined, hence the intelligence manifested by the program is a product of evolution, not of in-the-moment reasoning by the individual wasp. The individual wasp appears to be, like DAVE-2, running on autopilot, at least with respect to nesting behavior. This seems reasonable, given the wasp’s small brain and two-month adult lifespan. It has no time to learn the ropes, and nobody to learn them from, so it has to know how to reproduce and care for its young “right out of the box.”
Perhaps more interesting, though, is our own response to observing sphexish behavior. When we see the elaborate preparations the wasp makes for her brood, we think she’s smart and agential. Then, when we see that the behavior is pre-scripted, we “realize” that she’s just a machine, an “it,” and any impression of intelligence or willed action vanishes. But the difference is in our own minds, not that of the wasp.
Alan Turing arrived at a similar conclusion: “From the outside, […] a thing could look intelligent as long as one had not yet found out all its rules of behavior. Accordingly, for a machine to seem intelligent, at least some details of its internal workings must remain unknown.” 21
In the case of Sphex, this realization comes about due to our reverse-engineering of the script and “hacking” it to create an infinite loop. We come to believe the wasp has no intelligence or agency precisely because we can now predict its actions perfectly. What we really mean by “it’s just following a script” is that we, on the outside, know what the script is, and don’t even need to interact with the animal to predict what it will do next.
As a fellow computational being, how can you escape the same pickle—how do you keep from becoming a robotic “it” to a clever observer? Here are three ways:
- As described for moth flight in chapter 3, simply using a random variable can keep your behavior fresh and surprising even if you’re not so smart. In an adversarial setting, this strategy still works if your opponent is smart enough to see right through you (that is, if your internal state is transparent, or can be fully modeled by an outside observer). It even works if you are entirely generic and incapable of individual learning, meaning that your behavioral repertoire is fully genetically programmed, and you are “just like” every other member of your species. 22 Randomness is equally valuable in cooperative contexts, such as for foraging insect colonies, where it ensures that even near-identical individuals will diverge to explore different parts of their environment.
- The ability to learn allows your responses to be more unique, and allows your behavioral repertoire to scale up far beyond what can be encoded in your genome. By storing in your brain learnings and behaviors based on past experiences, you become vastly more complex on the inside—and all of this complexity is a hidden state, invisible to a stranger. In a competitive setting, such lifelong learning makes you harder to predict. In a cooperative setting, it allows individually differentiated skills and expertise to develop, greatly expanding the opportunities for cognitive division of labor.
- If you are able to model others effectively, including others who are modeling you back, then in an adversarial situation, you can outfox them by going meta. Modeling others is also essential to the cognitive division of labor, as it allows you to know who knows what you don’t know. Psychological studies suggest that we humans are so adept at keeping track of socially distributed knowledge that we tend to harbor greatly exaggerated beliefs about how much we personally know, when a good deal of our “knowledge” really consists of knowing whom to ask. 23
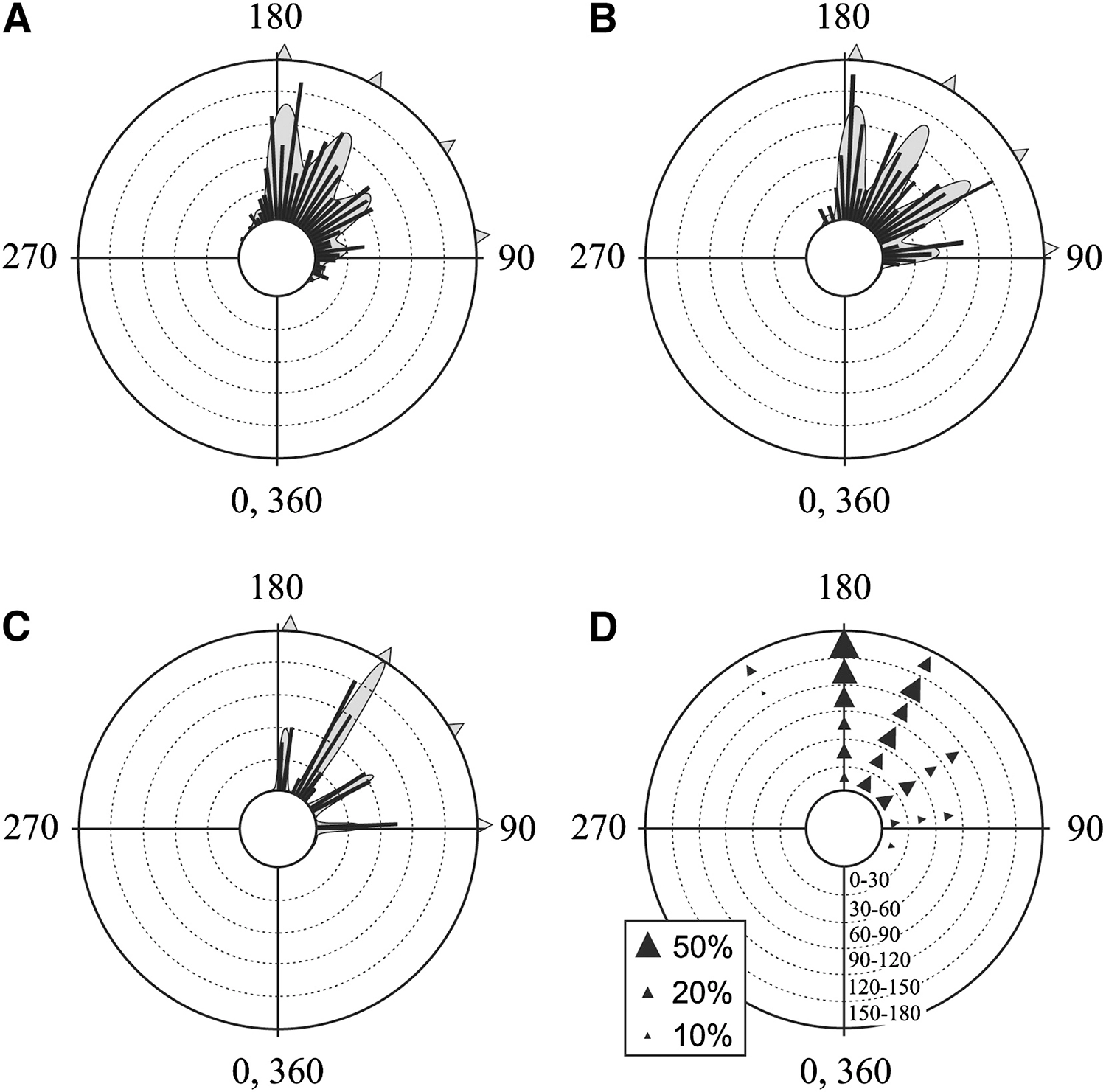
Polar plots of the escape trajectories of cockroaches startled by a gust of wind, showing random selection from among several preferred directions: (A) 431 pooled responses from five individuals; (B) frequency histograms of eighty-six individuals; (C) a histogram of the wind angles for the thirty-eight responses in (A) where the cockroach did not turn its body; and (D) a diagram illustrating the effect of wind direction on escape trajectory; for details, see Domenici et al. 2008.
Wasps might not bother with any of these strategies when it comes to nest preparation because cricket-moving tricksters have never exerted evolutionary pressure on making this specific behavior unpredictable. Predator/prey interactions are a different story. It’s unlikely that Sphex would behave so sphexishly while hunting the cricket it brings to its nest, or evading a predator of its own.
The last trick on the list—modeling your modeler—requires being very smart indeed. Roughly speaking, you need to be as smart as whomever you’re modeling, or more so, and you need to be able to imagine how you (and the world in general) appear from their point of view.
Does the small brain size of an arthropod preclude this trick? Apparently not. When Portia spiders show an awareness of what their adversary does and doesn’t know, and learn to actively manipulate that opponent’s world model, they are operating at this higher cognitive level.
Matryoshka Dolls
Among human children, this so-called “theory of mind” or ability to “mentalize” (I’ll use the terms interchangeably) takes quite a while to develop fully. The classic experimental setup for probing it is the Sally-Anne Test, popularized in 1985 by psychologists Simon Baron-Cohen, Alan Leslie, and Uta Frith. 24
The experimenters put on a little chamber drama for their young subjects using two dolls, Sally and Anne. Sally has a covered basket, and Anne has a box with a lid. Sally puts her marble in her basket, then goes out for a walk. While she’s gone, Anne slips the marble out of Sally’s basket and into her own box, closing the lid. Later, with Anne out of the picture, Sally returns.
The Sally-Anne Test being administered to a child
During the skit, the children are asked four questions:
- The Naming Question, a warm-up, asks them which doll is which; it establishes basic understanding of the scenario.
- On Sally’s return, the Belief Question is: “Where will Sally look for her marble?”
- This is followed up with the Reality Question: “Where is the marble really?”
- … and the Memory Question: “Where was the marble in the beginning?”
Even babies (by the time they can talk a little) are able to say who is who and understand where the marble is throughout the skit, although it’s stashed out of sight; hence questions 1, 3, and 4 are all fairly easy. However, until somewhere between two and a half and four years of age, children tend to get the Belief Question wrong. 25 They probably aren’t able to model the minds of others well enough to knowingly attribute false beliefs to them.
Psychologists tend to be very particular about which kinds of agents they believe could, even hypothetically, exhibit theory of mind, in part because of the baggage implied by language like “knowingly attribute.” Many reserve the term for humans only, though other great apes have also been shown to pass the Sally-Anne Test, with suitable modifications. 26 However, from a functional perspective, theory of mind is exactly what is required to lie or deceive, as Portia does (and as many other animals do). 27
Exhibiting this cognitive capacity implies not only an ability to simulate other minds, but also to model counterfactual (“what if”) universes. In the Sally-Anne Test, there is a real-life universe, in which the marble is in the box. There is the universe in Sally’s head, in which the marble is in the basket. Each of these universes also contains agents, like Sally and Anne, who in turn contain model universes in their virtual heads. The Anne in the universe in Sally’s mind, for instance, also believes that the marble is in the basket. Of course Sally and Anne are mere dolls; they only exist within the minds of the experimenter and the subject. The experimenter and subject are in turn abstract people I am describing to you, dear reader. Nesting dolls, universes within universes, in an endless recursion. It’s a dizzying prospect.
As adults, though, we negotiate such recursive mentalization all the time without a second thought. Reading a novel like Jane Eyre involves regularly entertaining fourth-, fifth-, and sixth-order theories of mind, as we wonder what Charlotte Brontë expects us to believe about what Jane believes Mr. Rochester must think Jane thinks about Mr. Rochester’s feelings about Blanche. Exercising this mentalizing faculty seems to have been the main pastime of the nineteenth-century English upper classes. (Writing popular novels about it added yet another layer of indirection, at times ironic.)
But theory of mind is far more than just a valuable skill for navigating hidden barbs and avoiding faux pas at teatime. There is good evidence that it is the very stuff of intelligence; it is, thus, at the heart of this book’s main argument.
We have arrived at that heart. My contention is that theory of mind:
- Powers the “intelligence explosions” observed in our own lineage, the hominins, and in other brainy species;
- Gives us the ability to entertain counterfactual “what-ifs”;
- Motivates, and is enhanced by, the development of language;
- Allows us to make purposive decisions beyond “autopilot mode”;
- Underwrites free will;
- Operates both in social networks and within individual brains;
- Results automatically from symbioses among predictors; and
- Is the origin and mechanism of consciousness.
In a sense, theory of mind is mind.
Intelligence Explosion
These are big claims. Let’s begin with the more established ones.
In the 1970s, Dian Fossey, the world’s leading expert on gorilla behavior, invited British neuropsychologist Nicholas Humphrey to spend a few months at her research station in the Virunga Mountains of Rwanda. Reflecting later on what he had seen, Humphrey wrote, “[O]f all the animals in the forest the gorillas seemed to lead much the simplest existence—food abundant and easy to harvest (provided they knew where to find it), few if any predators (provided they knew how to avoid them) … little to do in fact (and little done) but eat, sleep and play. And the same is arguably true for natural man.” 28
Archival footage from a 1973 National Geographic film documenting Dian Fossey’s gorilla studies in Central Africa, 1967–1972
These observations flew in the face of the usual explanation for evolving high intelligence—that it’s all about being a brilliant hunter, or otherwise “winning” at playing a brutal, Hobbesian game of survival in a tough environment. But if not to hunt (or evade hunters), why bother with intelligence? Couldn’t one live the easy life of a gorilla without incurring the high cost of a big brain? The fossil record suggests not: primate brains in many lineages, including those of gorillas and humans, haven’t shrunk over time, but, rather, have grown dramatically.
Humphrey’s explanation: “[T]he life of the great apes and man […] depend[s] critically on the possession of wide factual knowledge of practical technique and the nature of the habitat. Such knowledge can only be acquired in the context of a social community […] which provides both a medium for the cultural transmission of information and a protective environment in which individual learning can occur. […][T]he chief role of creative intellect is to hold society together.” 29
There are now many variations on this basic theory—some, like Humphrey, emphasizing cooperation and division of labor, others competition and Machiavellian politics. 30 These correspond fairly well to the “love” versus “war” explanations for the emergence of intelligence generally, as described in chapter 3.
In the 1990s, evolutionary psychologist Robin Dunbar and colleagues used comparative brain-size measurements across species to advance the closely related “social brain hypothesis,” which holds that the rapid increases in brain size evident in hominins and cetaceans (whales and dolphins), among others, arise from mentalizing one-upmanship. 31
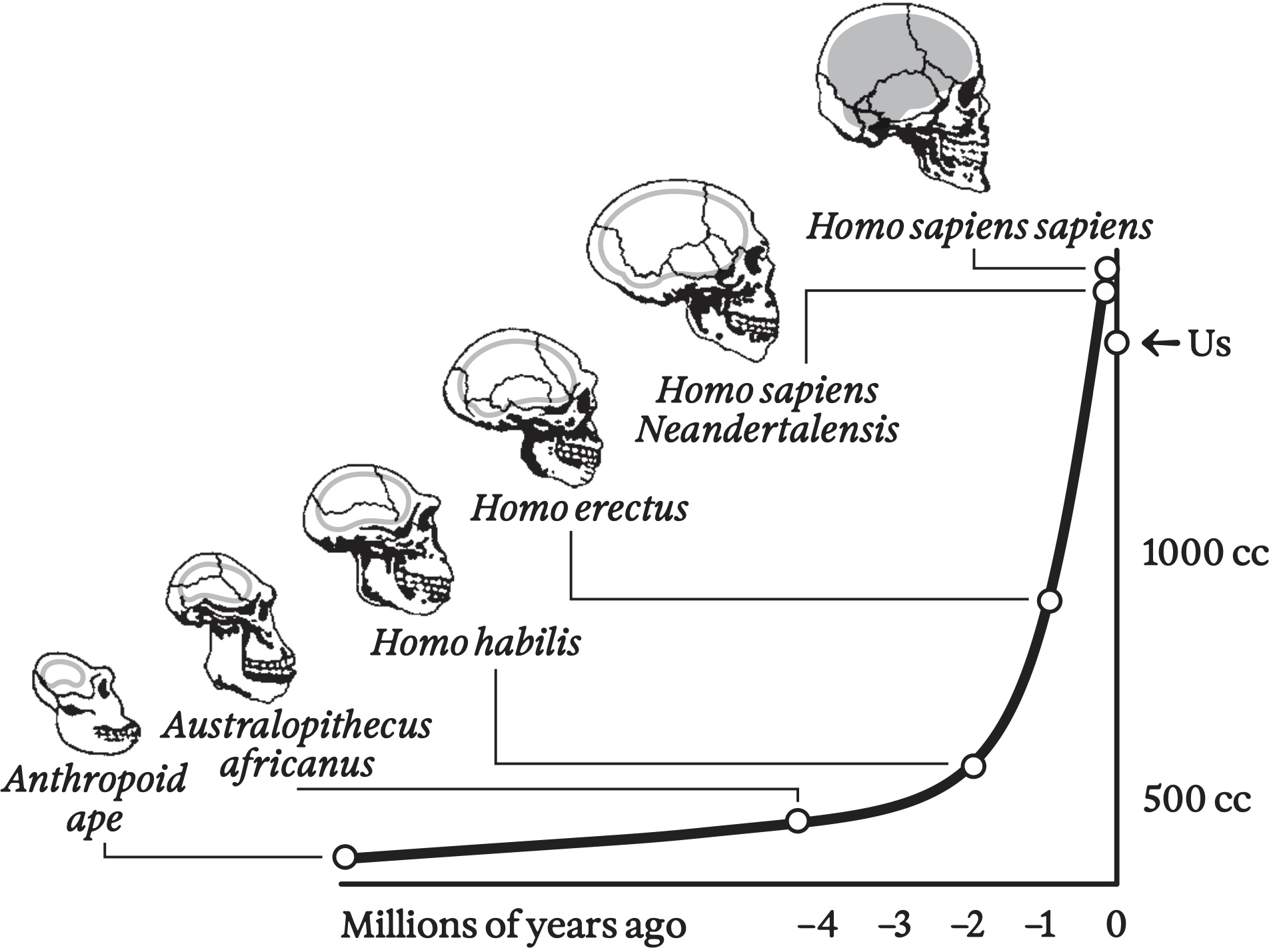
Explosion in hominin brain size over the past several million years (however, note the recent drop, possibly associated with the takeoff of social evolution and the division of labor)
Underpinning all of these related hypotheses is the observation that, among highly social animals like us, theory of mind is a powerfully adaptive trait. Being better able to get inside others’ heads increases the odds of finding a mate, building coalitions, securing resources from friends and family, getting help raising young, 32 avoiding violence (or being on the winning side of it), climbing in prestige, and amassing fans or followers. So, unsurprisingly, people with better theory of mind tend to live longer and have greater reproductive success. 33 That means Darwinian selection will be at work.
Strong theory of mind is correlated with both larger numbers of friends and larger brains, particularly in brain areas associated with mentalizing—above all, the frontal cortex. 34 Thus, startlingly, we can see evidence of the evolutionary pressure on social intelligence even among modern humans.
My guess is that the extra cortical volume of highly social people is dedicated not only to general mentalizing skills, but also to rich learned representations—we could even call them simulations—of their many specific family members, friends, colleagues, and acquaintances. 35 After all, mentalizing requires not just imagining another person in the abstract, but modeling their particular life experience, what they know and don’t know, their quirks and values, the ways they express themselves—in short, everything that comprises a personality, outlook, and umwelt. You probably bring such knowledge to bear effortlessly to play out hypothetical social situations in your head, drawing on a cast of hundreds of people you know.
This task is even harder than it appears at first glance, because there are infinite reflections in the social hall of mirrors: all of those people are themselves modeling others, including you. And, of course, their models of you include models of them, and of others. These relationships are important, because they powerfully affect behavior. Who shared their meat with whom after the last hunt, and in front of whom? Who is sleeping with whom? Who is gossiping about it, and who doesn’t know? Who is beefing with whom, and over what (or whom)?
Even if your model of second-order relationships (i.e., who is friends with whom, and what you know about those interactions) is not as rich as your model of first-order relationships (your friends), the sheer number of higher-order terms in your model explodes. If you have twenty classmates who all know each other (and you), then you need to keep track not only of your twenty relationships with them, but of all of their relationships with each other and with you, which is another 20×20=400 pieces of information. Third-order relationships climb into the thousands.
The numbers get truly mind-boggling when you consider that our acquaintances can easily number in the hundreds; family, school, and work environments tend to involve cohorts of people who all know each other; and our theories of mind can go up to sixth order, or beyond. 36 You can imagine, then, that even if many corners are cut, the amount of brain volume needed to do social modeling might grow both as a function of your number of friends and as a function of your ability to model higher-order relationships.
Indeed, when Dunbar and colleagues set out to find the relationship between the brain sizes and the social-group sizes of brainy animals, they found that as troop size increases, the amount of brain volume dedicated to the cortex also increases. 37 Mentalizing order, which Dunbar refers to as “intentionality level,” appears to be limited by cortical volume; behavioral studies suggest that monkeys can only operate at level one, while nonhuman apes have level-two intentionality. By extrapolation, archaic humans and Neanderthals may only have been able to achieve level-four intentionality, which is at “the lower end of the normal distribution for modern human adults, and at about the same intellectual level as young teenagers.” 38 Finally, the slope of the relationship between cortical volume and troop size is considerably steeper for apes than for monkeys, consistent with the idea that modeling higher levels of intentionality requires a greater investment of cognitive resources per troop member.
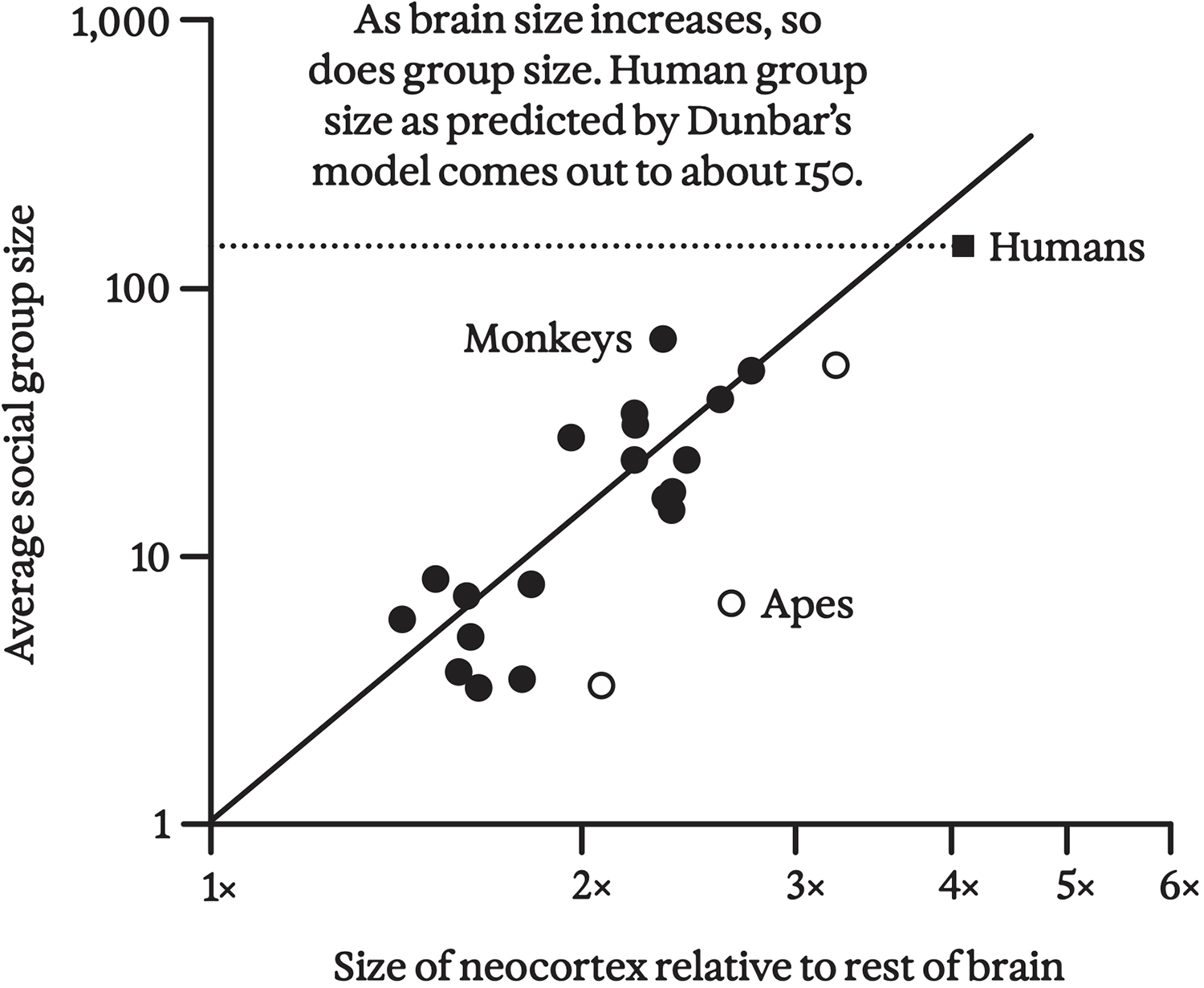
Correlation between neocortex size and social group size among monkeys and apes, redrawn from Dunbar 1998
Findings relating brain size to social group size, and social group size to Darwinian fitness, are themselves a hall of mirrors, revealing a profound self-similarity—and feedback loop—between brains and social groups. If you have a slightly larger brain than your friends and family, and are able to model more relationships more reliably, you will have a Darwinian advantage, so on average will have slightly more descendants than those who are less socially adept. But that means that your descendants will themselves become harder to model socially; that is, everyone else’s model of them must now be more complex, including higher-level intentionality and more relationships. And remember, everybody is trying to predict everybody else, but not be fully predictable themselves!
So it’s an arms race, not unlike that of the Cambrian explosion—though (usually) friendlier. Everyone is getting a bigger brain to model everyone else, and everyone else is getting harder to model at the same time, because … well, their brains are getting bigger. A social intelligence explosion ensues: a rapid (by evolutionary standards) increase in brain volume in your species.
When social modeling becomes such an important component of everyone’s life, the effect on individual incentives is dramatic. Lone operators, like leopards, are content to fend for themselves; their umwelt consists mainly of their territory and their prey, while other leopards are, most of the time, unwelcome intruders. For a modern human, though, being cast out or excluded from the community becomes a severe punishment—or even a death sentence. Our world consists largely of other people, and most of us would be unable to survive at all without continual mutual aid.
At the same time, sociality is a fraught business. We try to “win” at modeling others without being fully modeled ourselves; we compete for mating opportunities and for attention; we strive for dominance and prestige. These dynamics once again illustrate how competition and cooperation can be interwoven to such a degree that it can be hard to tell which is which.
Selection pressure also operates at the level of social groups. If one group sports slightly bigger brains and greater social prowess, the group itself can grow larger, and will thus tend to outcompete the smaller (and smaller-brained) group. Such may have been the fate of certain of our now-extinct hominin kin.
Societies have a kind of collective intelligence, and a rich body of work in social anthropology tells us that collective intelligence exhibits a scaling law not unlike that of individual brains. Theory of mind, incidentally, is important both for effective teaching and effective learning, which implies that it enhances cultural evolution at the group level too. 39 So, bigger societies can create and evolve more complex technologies, and thereby develop greater adaptability and resilience. 40 Greater scale, in other words, can support greater intelligence, and greater intelligence improves dynamic stability, both at the individual level and at the group level.
Crew of Eight
It’s hard not to wonder whether this recursive pattern applies at finer scales, too. Could our brains be “societies” of neural circuits that have evolved to model each other and thereby create the larger collective intelligence we call a “brain”? Neuroscientists and AI researchers have advanced such theories many times over the years, offering a range of supporting evidence. 41
For one, the cerebral cortex has a modular or repetitive structure, consisting of a honeycomb of “cortical columns”—though these columns don’t have distinct borders, which has led to endless debate about their exact definition and granularity. 42 Therefore, when I use the term “cortical column,” please take it with a grain of salt; I will interchangeably use even vaguer language like “small piece of cortex” or “part of the brain.”
Despite these ambiguities, we know that the basic “cortical circuit” is much the same in different regions of the cortex. When we distinguish between “auditory cortex” and “visual cortex,” for instance, the difference seems to be mostly about where each gets its inputs. 43 In one famous experiment, the optic nerves of baby ferrets were rerouted surgically to their auditory cortex, and these animals nonetheless developed the ability to see. Moreover, the characteristically patterned neural mapping of oriented visual features Hubel and Wiesel discovered in the visual cortex developed in the ferrets’ repurposed “auditory” cortex, albeit more sloppily. 44 There’s evidence that the human brain can rewire in similarly dramatic ways: blind people can learn to see using their hearing via “click sonar,” 45 and can even acquire a limited form of vision via spatially patterned tongue stimulation. 46
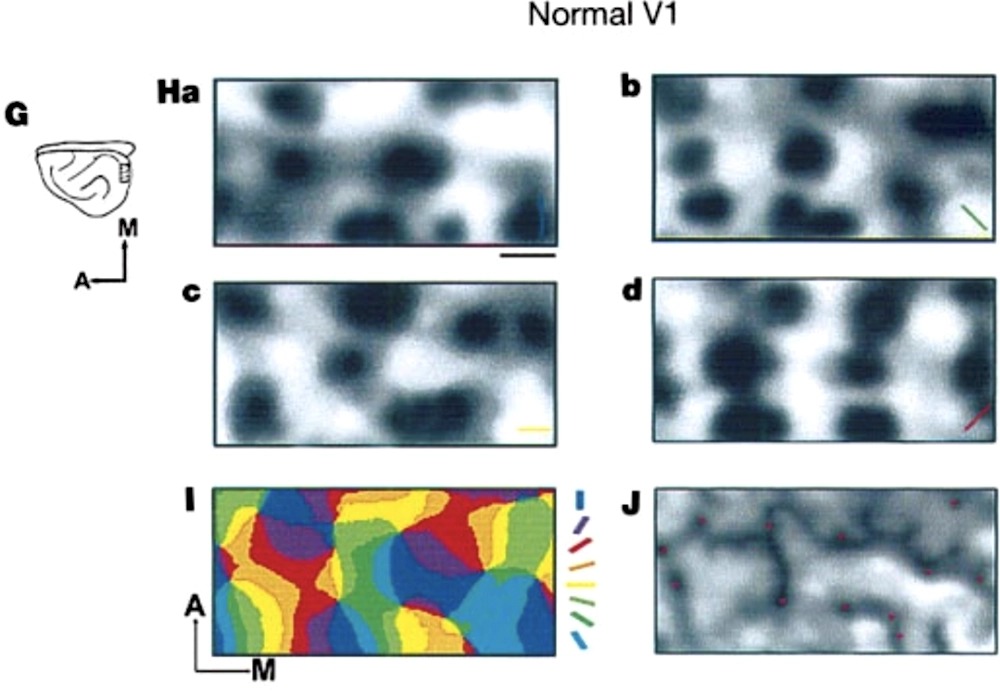
Maps of visual stimulus orientation sensitivity in a normal ferret’s visual cortex (V1), followed by a similar map in the auditory cortex (A1) for a ferret whose optic nerves have been “rewired” to feed into its auditory cortex. Darker regions represent cortical locations with higher activity, and colors show the angle of the visual grating producing the strongest response; Sharma, Angelucci, and Sur 2000.
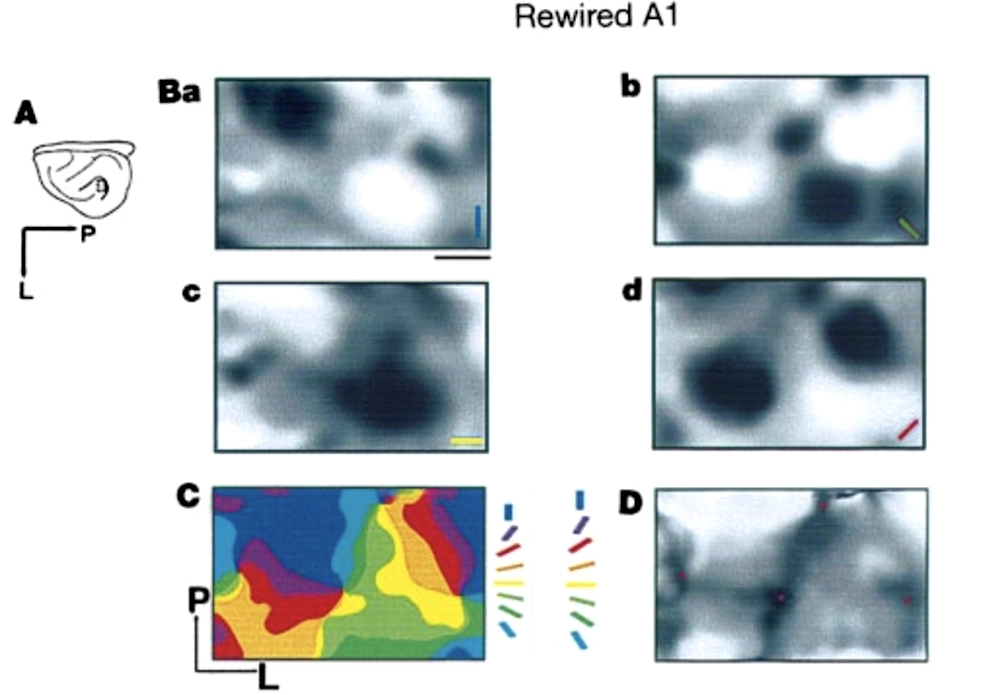
Maps of visual stimulus orientation sensitivity in a normal ferret’s visual cortex (V1), followed by a similar map in the auditory cortex (A1) for a ferret whose optic nerves have been “rewired” to feed into its auditory cortex. Darker regions represent cortical locations with higher activity, and colors show the angle of the visual grating producing the strongest response; Sharma, Angelucci, and Sur 2000.
Perhaps generic modularity is precisely what made the intelligence explosion possible: just as DNA could easily evolve to replicate more vertebrae and ribs to produce snakes (per chapter 1), the genomes of animals undergoing intelligence explosions could respond to individual and group selection pressure by replicating more cortical columns. Thus, evolution could progressively expand the cortical sheet without needing to invent anything fundamentally new. In the biggest-brained animals, like dolphins and humans, expansion progressed to the point of scrunching up the cortex into dense folds, giving our brains their characteristic furrowed appearance. We manage to cram about a quarter of a square meter of cortical area into our skulls.
Given this, one might even consider the cortex itself to be more like a population than a single entity: a “colony” of cortical columns that have managed to replicate inside our skulls in larger and larger numbers through increased cooperation among themselves. The greater intelligence they give us adds to our individual fitness, which in turn allows us to form larger societies, which increases dynamic stability all round.
You might find it disconcerting to think of your cortex as a colony of entities, rather than as one unified entity, “yourself.” Remember, though, that at some level, life always works this way. Our cells harbor colonies of mitochondria, and our bodies consist of colonies of cells. All of these small entities are intelligent in their own right, as any living thing needs to be in order to survive when surrounded by other similar entities in a dynamic environment.
I think the reason we find intelligent cortical columns uncanny is that their umwelt, the domain in which they are intelligent, overlaps with “our” umwelt—unlike, say, a white blood cell, which, while obviously alive and active, inhabits a microscopic inner world utterly alien to us. Cortical columns, by contrast, understand colors and shapes, people and places, stories and emotions. Our understanding is their understanding. And that is weird.
Indulge me in a speculative digression about everyone’s favorite “alien intelligence”: the octopus. 47 Octopuses have always seemed like a bit of a fly in the ointment, as far as the social-intelligence hypothesis is concerned. Unlike other very smart animals on Earth, they’re notoriously cranky and individualistic. They don’t tend to hang out with each other, and usually only meet to mate—a fraught encounter that may end in violence or even cannibalism. 48
Octupuses mating
While they are brilliant lifelong learners, they don’t live long enough to raise their young, hence lack the ability to accumulate cultural knowledge. If they did, it seems entirely possible that our planet’s seas would teem with octopus cities, and perhaps even octopus-made computing machines (which would undoubtedly use an octal, or base eight, numbering system—a much better engineering choice than our own base ten).
If octopuses are so antisocial, and intelligence is a social phenomenon, why are they so smart? One could dodge the question and note that sociality doesn’t just involve conspecifics (i.e., others of your own species), nor is it necessarily friendly. With omnivorous appetites and a total lack of body armor, the octopus must rely on its wits to model and predict everything it wants to either hunt or escape from; it seems likely that highly developed theory of mind, vulnerability to predators, and flexible hunting strategies all coevolved. 49
A more fun possibility, though, is suggested by considering the other ways octopuses are so different from other big-brained animals, including us. The octopus’s “brain” is highly decentralized, likely due to the high cost of long-distance communication in the nervous systems of molluscs. Our own long-distance nerve fibers are coated with fatty myelin sheaths, which greatly speed up and lower the energetic cost of transmitting electrical impulses. 50 Mollusc nerve fibers lack myelin sheaths, though. Transmitting fast impulses in unmyelinated nerves requires dramatically increasing the diameter of the axon, which explains why squids have a “giant axon”—a single nerve fiber up to 1.5 millimeters in diameter!—running the length of their body. This giant axon allows for a rapid, coordinated escape response. 51 But obviously, for an animal with many neurons powering complex behaviors, there isn’t room (or the energetic budget) for many such giant axons, so a great majority of neural processing needs to be local. That probably explains why three fifths of an octopus’s neurons are in the arms, rather than in the head.
While cephalopods are bilaterian like us, their bodies are far more modular than any land animal, exhibiting many additional near-symmetries. In the octopus, over two hundred suckers are arranged in beautiful patterns along each of eight arms, with chained neural networks surrounding each sucker. Individual suckers are prehensile, almost like a pair of opposed fingers, and endowed with far richer local sensory input than our hands, including touch and taste receptors, chromatophores for colorful skin displays, and even photoreceptors for light sensing. A lot of tasting, moving, color-changing, reacting, deciding, predicting, and even reasoning seems to occur under local control.
Extreme modularity of the octopus body; not only is each arm intelligent, but so is each sucker
Hence the octopus’s modularity is not just structural, but functional. Each sucker is smart. Each arm, too, is intelligent, able to respond to stimuli with complex movements without the involvement of the central brain. 52 Arms recognize each other directly as they move, avoiding self-entanglement without requiring any central representation of the animal’s geometry, 53 which—given the wild range of geometries possible for such a squishy animal with so many degrees of freedom—would be an extraordinary challenge for any brain. 54 The arms can even communicate with each other directly via a ring of ganglia that encircle the “shoulders,” with additional direct connections between alternating arms; these routes bypass the brain entirely. 55
There’s more. Octopuses occasionally need to bite off an injured arm, an act known as “autotomy.” 56 They may even do so to distract a predator, leaving one writhing appendage behind while the other seven, along with the head, escape. (At this point, as a smartass on Reddit observes, we would have a septopus.) In at least one squid species, “attack autotomy” has been observed as well: a severed arm attacks a predator while the rest of the animal escapes. 57 Following autotomy, a new arm will regrow. For octopuses, the process takes about 130 days.
Partial autotomy in the squid Octopoteuthis deletron when its arms get stuck in a bottle brush. Note the continued activity of the severed arm tip; Bush 2012.
Perhaps, then, the octopus is best thought of as a tightly knit community of eight arms, sharing among themselves a common pair of eyes. In fact, the bulk of the central “brain” consists of a pair of optic lobes; we could think of these lobes compressing visual information in service to the arms, rather than being “the boss,” or even, in any meaningful sense, “the octopus.” In this light, attack autotomy in the squid (whose nervous system is similarly organized) may not be so different from the kamikaze tactics of a bee defending the hive.
Could the intelligence explosion that resulted in the octopus have been driven by predictive social modeling—theory of mind, in effect—among its eight arms? As absurd as this sounds, I think it’s plausible. Octopus arms are individually smart, but communication among them is limited, due both to the high latency and low bandwidth of unmyelinated nerve fibers. Centralized control under these constraints is impossible.
Yet the octopus’s arms must work together seamlessly to swim, hunt, escape aquarium tanks, and so on; to do so, they must model each other. We know that people can work closely together to perform such tasks with only a little communication; consider an elite military unit, or a hunting party, exchanging subtle signs while stalking their quarry. Even intellectual tasks, like solving escape-room puzzles or competing in Math Olympiads, involve such teamwork. Language is a very low bandwidth medium, yet we still manage.
Accounts of small, extremely high-performing teams invariably emphasize both the unity of purpose that can be achieved and the need for each team member to infer what the others are up to based on minimal cues: that is, high-fidelity mutual prediction, or mentalizing. When this happens in competitive rowing, it’s called “swing”: “It only happens when all eight oarsmen are rowing in such perfect unison that no single action by any one is out of sync with those of all the others. […] Each minute action […] must be mirrored exactly by each oarsman, from one end of the boat to the other. […] Only then will it feel as if the boat is a part of each of them, moving as if on its own.” 58
Archival footage from the 1936 Olympics in Berlin, where the scrappy University of Washington rowing crew won a gold medal; Brown 2013.
Homunculus
How different are we from the octopus, really? We have fast, myelinated nerves that allow for greater physical centralization of our brain in our heads, but if what I’ve suggested is correct—that our brains can be thought of as symbiotic colonies of cortical columns modeling each other—then the difference may be more of degree than of kind. This is unsettling, because we tend to have a strong sense of ourselves as a unified “I.”
Much intuitive (and supernatural) thinking over the ages has presumed this indivisible self, located somewhere in the body. Presumably it’s in the brain somewhere. That’s why we can imagine getting a “full body transplant” but not a “brain transplant.” A common term for this “something in the brain” is a “homunculus,” literally, a “little man” somewhere in there, pulling the strings. In many religious traditions, the immortal soul plays this homuncular role.
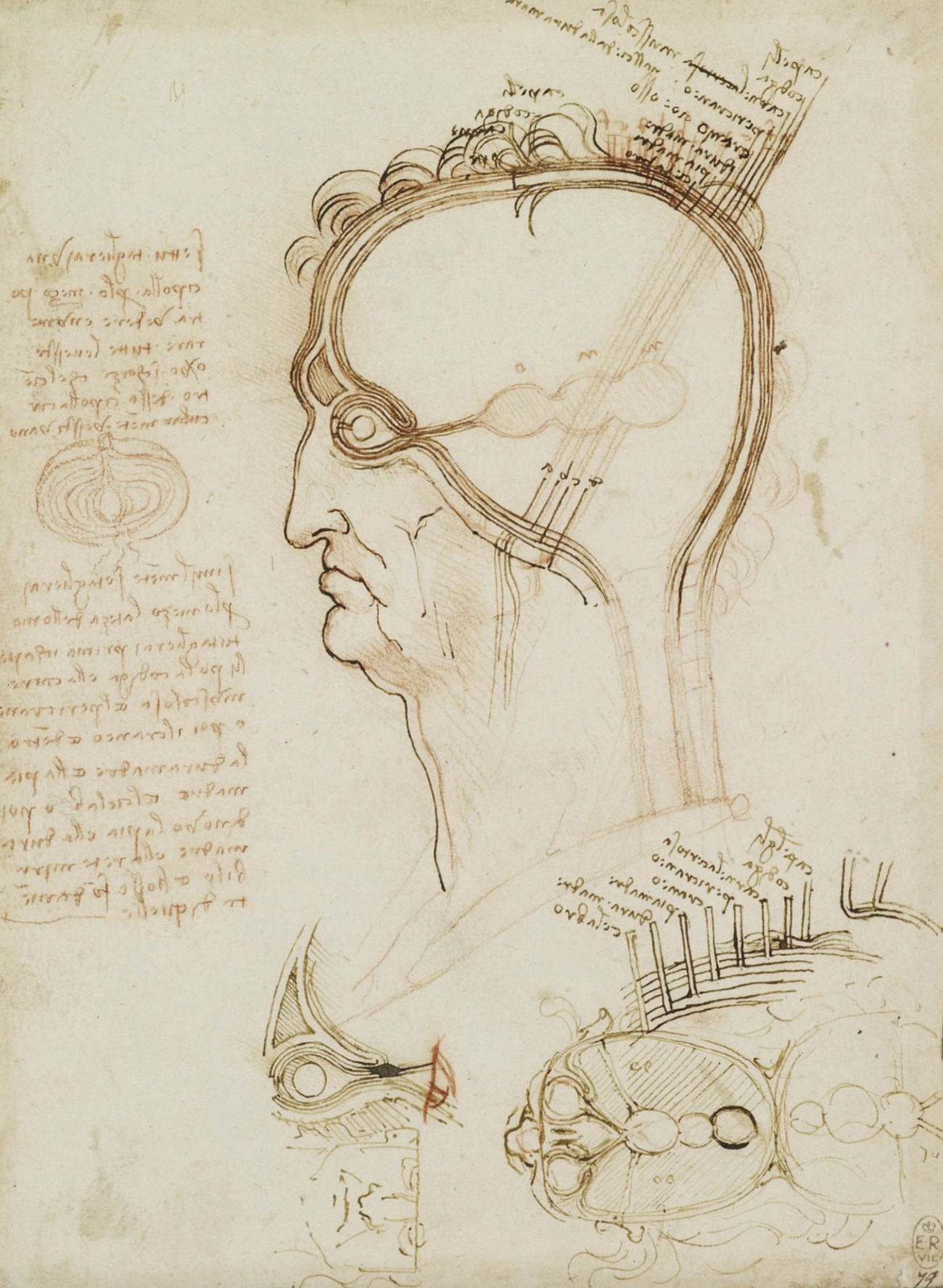
Leonardo da Vinci’s early understanding of neuroscience was largely based on received wisdom from Galen about the cognitive functions of three ventricles corresponding to imagination, reason, and memory. By 1509, da Vinci had carried out extensive dissections and experiments, including the use of wax to make moulds of the cerebral ventricles of an ox, resulting in groundbreaking anatomical accuracy but, as yet, little conceptual progress.
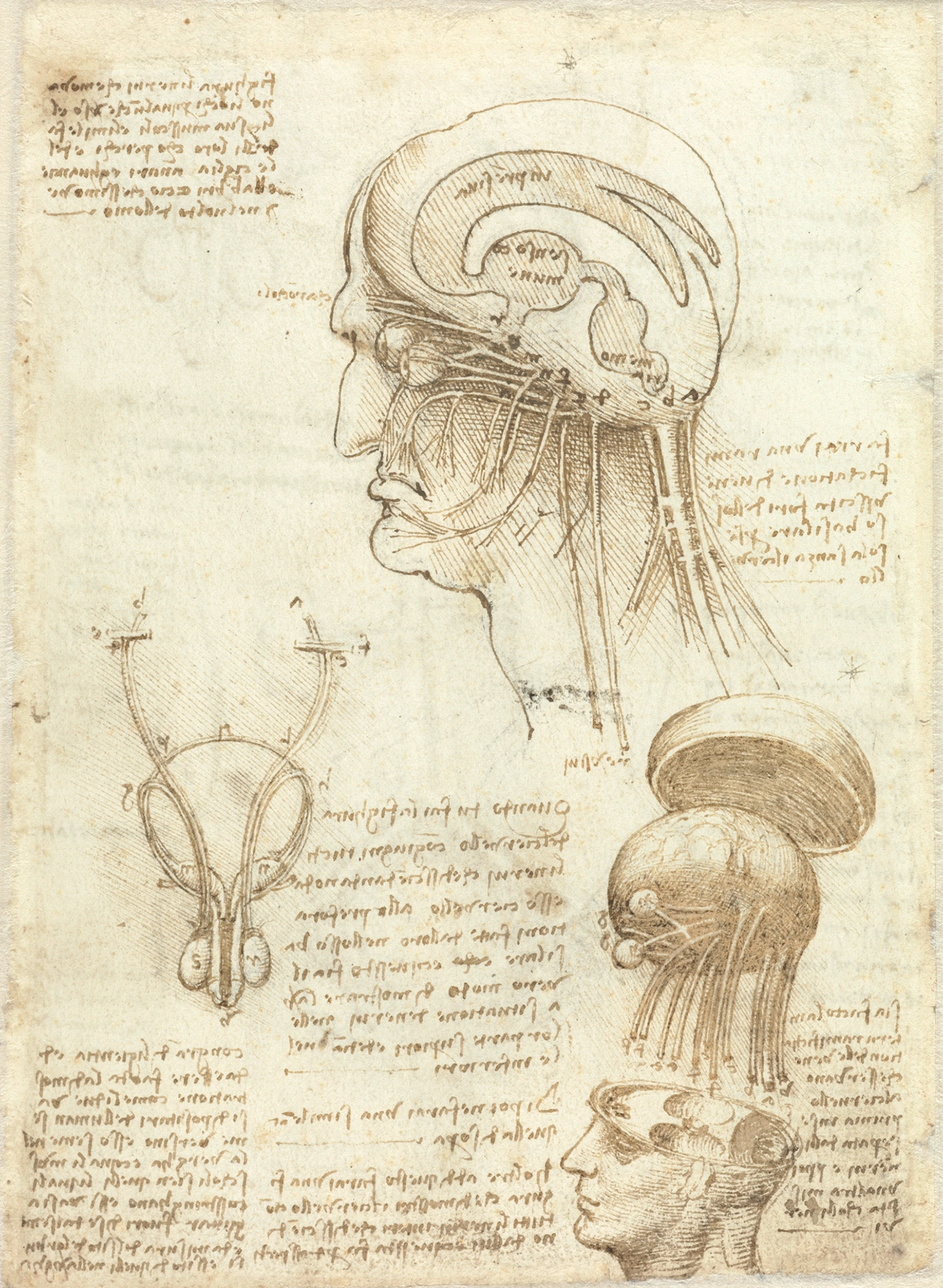
Leonardo da Vinci’s early understanding of neuroscience was largely based on received wisdom from Galen about the cognitive functions of three ventricles corresponding to imagination, reason, and memory. By 1509, da Vinci had carried out extensive dissections and experiments, including the use of wax to make moulds of the cerebral ventricles of an ox, resulting in groundbreaking anatomical accuracy but, as yet, little conceptual progress.
René Descartes (1596–1650) thought about the problem from a neuroanatomical point of view. An influential champion of “mechanical philosophy,” or what we now call physics, Descartes held that the body and brain must be mechanical, or machine-like, since we seem to be made of the same stuff as the rest of the physical universe. Further, human anatomy has much in common with other animals’ anatomy, and human behavior has much in common with other animals’ behavior: the “passions” of hunger and sexual attraction, reflexes to flinch if burned, and so on. Hence Descartes coined the term “bête-machine,” or “animal-machine,” to describe the mechanically based physiology of animal and human bodies.
Still, Descartes needed some way to reconcile mechanical philosophy with his Christian faith. 59 The Church held that we have immortal souls while other animals do not, and the Enlightenment emphasized the divine gift of human rationality; other animals certainly didn’t seem to be writing philosophical treatises. Borrowing from Aristotle, Descartes found an elegant solution: identifying rationality itself with the soul. The twist was that a soul-brain interface of some kind would be needed for the immaterial soul to puppet the material body.
To theorize how this might work, Descartes again borrowed from the classical world, this time a medical authority. Galen (roughly 130–210 CE) believed the cerebral ventricles were filled with “psychic pneuma,” a volatile, vaporous substance that he described as “the first instrument of the soul.” 60 Riffing on this idea, Descartes imagined the body to be, literally, a hydraulic machine powered by psychic pneuma: “When a rational soul is present in this machine it will have its principal seat in the brain and will reside there like the fountaineer, who must be stationed at the tanks to which the fountain’s pipes return if he wants to initiate, impede, or […] alter their movements.” 61
Clever! But where was this “fountaineer”? Descartes concluded that it must reside in the pineal gland, a small, pinecone-shaped structure near the center of the brain. For one thing, it’s near the cerebral ventricles, the supposed “tanks” or reservoirs of psychic pneuma. More importantly, the pineal gland is one of the few discrete structures located along the brain’s midline—meaning we have only one of them. Being indivisible, surely the soul couldn’t reside in a pair of structures on both the left and right sides of the brain!
Illustrations of the pineal gland, seat of the soul, hydraulically “puppeteering” the body using psychic pneuma; Descartes 1677
Spoiler alert: the pineal gland’s main job is melatonin production. It helps regulate the circadian rhythm. It is not the seat of the soul.
Descartes’s mistake was more fundamental than picking the wrong gland. Some neuroscientists are still seeking an elusive brain region, circuit, or type of neuron where our consciousness might reside. It’s certainly true that not all brain regions are created equal; the severity of a stroke depends critically on which structures it knocks out. However, I think that attempting to pin down the location of a homunculus, soul, or “self” within the brain is like trying to locate where the “swing” is in a crewed boat.
This isn’t to say that either phenomenon is unreal or illusory. Swing is perfectly real, as those who have experienced it can attest. But it is a verb, not a thing; distributed, not localized; a dynamic process in time, not a static condition; and relational, not individual. There is nothing supernatural about it, but neither is it physical, or even objective. It’s functional. It results in higher performance of the boat, and so has tangible effects in the world. But it is also subjective, experienced by each of the eight members of the crew in relation to the others, and to the whole.
Imagine replacing one of the crew members in the boat with a robot. Not the dumb kind we have in factories today, but one capable of pulling the oars like a person, hearing, seeing, touching, and using haptic and audiovisual cues to communicate with the rest of the crew. Could such a “cyborg crew” achieve swing? I think the answer is a clear “yes,” and, in fact, given the sophistication of today’s AI models, any remaining impediment has mainly to do with deficiencies in batteries and motors—bodies, not “brains.”
This thought experiment is a boat-crew version of one posed by philosophers David Chalmers and Susan Schneider that we could call the “brain of Theseus,” 62 by reference to Plutarch’s much older “ship of Theseus” thought experiment (to stick with the boating theme a little longer). Theseus, the mythical king and founder of Athens, has a legendary ship that is preserved by the Athenians for generations. It undergoes innumerable repairs over the years to keep it in good order. The question is: once every timber has been replaced, is it still the ship of Theseus?

Something close to a real-life ship of Theseus: the USS Constitution, built in 1797. The first photo, from 1858, shows it under repair at the Portsmouth Navy Yard; in the second photo, at the Charlestown Navy Yard in 1927, it is undergoing a three-year restoration involving the “renewal” of 85 percent of the ship; and finally, after a major round of repairs in 2015, little of the original wood remains.

Something close to a real-life ship of Theseus: the USS Constitution, built in 1797. The first photo, from 1858, shows it under repair at the Portsmouth Navy Yard; in the second photo, at the Charlestown Navy Yard in 1927, it is undergoing a three-year restoration involving the “renewal” of 85 percent of the ship; and finally, after a major round of repairs in 2015, little of the original wood remains.

Something close to a real-life ship of Theseus: the USS Constitution, built in 1797. The first photo, from 1858, shows it under repair at the Portsmouth Navy Yard; in the second photo, at the Charlestown Navy Yard in 1927, it is undergoing a three-year restoration involving the “renewal” of 85 percent of the ship; and finally, after a major round of repairs in 2015, little of the original wood remains.
Now that we understand something about cellular biochemistry, we know that the question applies literally to us, too, as most of the cells in our bodies turn over many times throughout our lives, and, even in the longest-lived neurons, every atom will get swapped out over time. If we want to assert the continuity of our own identity, we need to acknowledge that a “self” is, like swing, a dynamical and computational process, not a specific set of atoms. And the same is true of a ship!
Let’s follow Chalmers and Schneider in considering the “ship of Theseus” question for the brain, now with a cyborg twist, as for the rowing crew. Imagine that a single neuron in your brain is replaced by a computer model of that neuron. Once more, the greatest problems here are technological, not conceptual. Thanks to those mid-century experiments on the squid giant axon (and a great deal of work since), we have excellent computational models of individual neurons, at least with respect to their fast dynamics.
A complete neuron-machine interface is a much taller order. It would require not only coupling a computer to all pre- and post-synaptic neurons, but also the ability to sense and emit a variety of neuromodulatory chemicals. This is well beyond us today. But let’s set practicalities aside. The question is: if one of your neurons were replaced by a “robot neuron,” would you notice?
Of course you wouldn’t, because the brain is robust to anything going awry with any single neuron. But what if we repeated the procedure for a billion of your neurons? Or half of them? Or all of them?
The question is unsettling, because it highlights the difficulty we have in coming to grips with a dynamic, process-oriented, computational, distributed, and relationship-centric view of the “self.” We are still Cartesian enough in our thinking to wonder whether—per Schneider’s description of a hypothetical surgeon replacing your biological neurons with silicon-based ones—“your consciousness would gradually diminish as your neurons are replaced, sort of like when you turn down the volume on your music player. At some point […] your consciousness just fades out.” Or perhaps, “your consciousness would remain the same until at some point, it abruptly ends. In both cases, the result is the same: The lights go out.”
These ideas are—I have no words to put this gently—silly (and I think Schneider and Chalmers both agree). In physical and computational terms, a neuron has an inside and an outside, and if the insides were scooped out and replaced with different machinery that performed the same function—which is to say, resulted in identical interactions as seen from the outside—then none of the larger functions in which this neuron plays a part will be affected. Per the Church-Turing thesis, these functions could be computed in many equivalent ways, and using many different computational substrates. That is true at every level of the nested set of relationships, or functions, that we call “life” or “intelligence.”
Illusion and Reality
Philosopher of mind Daniel Dennett believed that consciousness and the self are illusions; we’re robots made of robots made of robots, so, per Dennett, there is no real “there there,” other than as a convenience of speech. 63 Similarly, for philosopher-neuroscientists Sam Harris and Robert Sapolsky, 64 free will is an illusion because minds are products of inexorable physical processes.

Illusionism, an anthology of academic articles about consciousness as an illusion, including contributions from Daniel Dennett and Nicholas Humphrey; Frankish 2017.
We are indeed robots made of robots made of robots, and minds are indeed based on physical processes. However, my own sense is that “illusion” isn’t necessarily a helpful word here, because it implies an incorrect assumption, or at best a polite fiction. While nobody has suggested that we dispense with this “fiction” altogether, Sapolsky comes close to running off that cliff when he asserts that, since minds are physical and free will is illusory, notions like criminal justice, prizes, praise, blame, and moral responsibility are all invalid and should be abandoned.
In Sapolsky’s defense, it is worthwhile to ask tough questions about our received notions of justice, reward, and responsibility, especially when confronted with alternatives that seem more humane, fairer, or lead to better outcomes. It’s less productive to become nihilistic because “it’s all an illusion.” That kind of nihilism could also lead us to conclude that tables and chairs aren’t “real,” 65 because they, too, are mere collections of atoms.
Chapter 2 offers a more pragmatic take on reality, familiar from physics as well as real life: “reality” is our name for a model with good predictive power. Or rather, a suite of models. We will always need many, because every model has a limited domain of validity. It would be awesome to discover the physicists’ holy grail, a “Theory of Everything” unifying quantum fields and gravity. But it would tell you nothing about whether your aunt will like the cake you’re baking for her, your friend will take your shift at the café, or your kid will get along with the neighbor’s kid. In fact, unless you’re one of a handful of rarefied physicists, it will inform precisely nothing in your life.
Good questions to ask about a model, then, are: Does it agree with observations? Does it make any testable predictions? Who cares about it? What function does it serve? Where does it break down? Do we have a better candidate model? Sometimes we do have a better candidate, but, as with general relativity, it may be disturbing or challenge deep-seated intuitions. Newtonian physics, which we find a lot more intuitive, follows from general relativity, but only as an approximation valid in our everyday world, where spacetime is reasonably flat and objects move far slower than the speed of light.
That’s not to say that classical physics is an “illusion”; rather, the more general theory illuminates a wider area. Einstein’s theory shows us when the Newtonian approximation holds, when it doesn’t, and why. It resolves apparent contradictions, like the incompatibility of Newtonian physics with a constant speed of light. In this sense, general relativity actually bolsters the narrower classical theory by reassuring us that the seeming paradoxes aren’t cause for concern, and by showing us why, when, and to what degree we can rely on familiar approximations.
Free will; the unitary, indivisible “self”; and interaction with other unitary “selves” in the world are the common-sense axioms underpinning something like a Newtonian model of social life. Theory of mind is that model. It’s our folk theory—literally. It’s what we use when we predict our aunt’s tastes, our friend’s willingness to help, or our kid’s friendships. While powerful and useful in everyday life, theory of mind is also confounding, even paradoxical, when we try to reconcile it with the physical universe; hence the trouble Descartes got us into. In the following discussion, we’ll flesh out a more general theory that will help us better understand our folk theory—and how it arises, and its conceptual limits.
But let’s keep in mind just how powerful that folk theory is. Will Aunt Millie like my cake? For a physicist, making such a prediction may seem like weak tea compared with the extraordinarily precise, quantitative predictions we expect from physical theories. But it’s not so.
The parable is named after the fourteenth-century French philosopher Jean Buridan, but versions of it long predate him.
The earliest versions of randomized or “Monte Carlo” methods (named after the famous casino) were implemented at Los Alamos by Augusta Teller and Arianna Rosenbluth to simulate chain reactions in nuclear weapons.
Silver et al. 2017 ↩.
Schrittwieser et al. 2020 ↩.
Silver et al. 2013 ↩.
Kahng and Lee 2016 ↩.
Churchland 2016 ↩.
It does, in an oblique way, maintain a memory of prior moves: the 19×19-pixel image used by AlphaGo’s convolutional nets to evaluate the board includes “channels” representing, for instance, the number of moves since the stone was placed.
My old college roommate used to hustle chess games in New York’s Washington Square Park. First, he and I would play for money; I was genuinely bad, but he only pretended to be. He would then clean up against the other chess hustlers, who would initially play sloppily, assuming he was an easy mark. Afterward, we would split the proceeds.
I’m simplifying slightly. AlphaGo was designed to “cache” Monte Carlo evaluations of board positions so that they could be reused without being recomputed during subsequent moves; however, this was an engineering optimization. It didn’t involve the introduction of hidden states that influenced gameplay.
R. R. Jackson and Pollard 1996; Harland and Jackson 2000 ↩, ↩.
Wilcox and Jackson 2002 ↩.
Tchaikovsky 2018 ↩.
Before you risk Googling it, recall from “Behavior, Purpose, and Teleology,” chapter 3, that an entity’s Wiener sausage is the tubular zone of uncertainty it maintains around its trajectory through the world, thus avoiding being fully predictable.
Chiang 2005 ↩.
Hofstadter 1982 ↩.
Chittka 2022 ↩.
Fabre 1916 ↩.
Insects exhibit meaningful individual differences in intelligence, just as humans do; when Fabre tried the trick on a different Sphex population of the same species, he found that they weren’t fooled, per Chittka 2022 ↩. We’ll return to the Sphex story with a more critical eye in “Hive Mind,” chapter 9.
Riskin 2016 ↩.
In reality, no living being can be truly identical with another, as even the simplest organism has a vast number of internal degrees of freedom, and many are neutral, i.e., confer neither a survival advantage nor disadvantage.
Sloman and Fernbach 2018 ↩.
Wimmer and Perner 1983; Baron-Cohen, Leslie, and Frith 1985 ↩, ↩.
Setoh, Scott, and Baillargeon 2016 ↩.
Krupenye et al. 2016 ↩.
Ding et al. 2015 ↩. Note that the spider may still exhibit social competence without comprehension—meaning that it may not model itself attributing false beliefs to others, which would amount to a higher-order theory of mind.
Humphrey 1976 ↩.
The basic idea had been floated by other researchers in the 1950s (Chance and Mead 1953 ↩) and again in the ’60s (Jolly 1966 ↩), though to less effect.
Dunbar 1998 ↩.
Sarah Blaffer Hrdy has convincingly argued that, among humans and certain other brainy species, it “takes a village” to raise an infant, in the sense that the mother alone can’t provide all the needed calories; the grandparents, siblings, and babysitters who help out are called “alloparents.” In alloparental species, prospective mothers without the needed social support disproportionately elect infanticide or, among humans, abortion. If they do have the baby, its odds of survival increase appreciably with alloparental help; Hrdy 2009 ↩.
Pawłowski, Lowen, and Dunbar 1998; Holt-Lunstad et al. 2015 ↩, ↩.
Powell et al. 2012 ↩.
There’s likely no sharp boundary between general skills and specific knowledge, in this or any other domain, because every specific thing you learn is represented in terms of your existing conceptual vocabulary, and in turn extends that vocabulary. One can think of it almost like a compression algorithm, in which one’s experiences so far make up the “dictionary” with which subsequent knowledge and experiences are compressed. This may (at least partly) explain why time seems to pass more quickly as we age: we’re compressing our life experiences more efficiently—alas.
Street et al. 2024 ↩.
Dunbar 1992 ↩.
Dunbar 2016 ↩.
Muthukrishna et al. 2014 ↩.
Mountcastle 1957; Buxhoeveden and Casanova 2002; Cadwell et al. 2019 ↩, ↩, ↩.
Buonomano and Merzenich 1998 ↩.
Sharma, Angelucci, and Sur 2000 ↩. Note that the claim has recently been challenged by some researchers, who argue that apparent instances of cortical remapping (especially after brain trauma) are illusory, and merely reflect greater use being made of pre-existing, latent capabilities in neighboring areas of cortex; T. R. Makin and Krakauer 2023 ↩. It is indeed likely that cortical maps with well-defined boundaries are somewhat of an illusion. However, to me, the evidence—both from neuroscience and from machine learning, where the same neural net architecture can learn to process very different inputs—suggests that cortical remapping is real, and explains how it could occur without requiring a radically altered architecture or learning algorithm.
Kolarik et al. 2014 ↩.
Chebat, Schneider, and Ptito 2020 ↩.
Godfrey-Smith 2016 ↩.
Exceptional sites have been found where groups of octopuses live together; Scheel et al. 2017 ↩.
Octopuses have also been around for a long time, and we have no way of knowing what their social behaviors might have looked like a hundred million years ago. Perhaps their intelligence evolved at a time when they were nicer to each other.
Myelination is what makes our longer nerve fibers whitish; hence the unmyelinated “gray matter” of the cerebral cortex, where connectivity is mostly local, and the “white matter” of the brain’s interior, which consists mostly of wiring.
Since its enormous size makes it so easy to work with in the lab, the squid giant axon played a starring role in the midcentury experiments that established the biophysics of the action potential.
Sivitilli and Gire 2019 ↩.
Crook and Walters 2014 ↩.
Since the beak is an octopus’s only rigid part, it can squeeze through tiny holes or cracks—anecdotally, pretty much any space big enough for one of its eyeballs to get through; Godfrey-Smith 2016 ↩.
Kuuspalu, Cody, and Hale 2022 ↩.
Budelmann 1998 ↩.
S. L. Bush 2012 ↩.
D. J. Brown 2013 ↩.
While Descartes’s faith was probably genuine, self-preservation undoubtedly played a role, too. As we shall see, taking the mechanical philosophy too far, even a century later, was dangerous.
Galen 1968 ↩.
Descartes 1677 ↩.
Tables and chairs seem to be invoked by philosophers more often than anything else when appealing to “reality,” presumably because they are so ubiquitous to a philosopher’s umwelt.